源容量
专注于生物质生产的限制因素,我们遇到了植物源器官中一次能量固定的可能限制的问题。当然能量的吸收光吸收不是一个限制因素,因为光合作用固定在中等光子通量密度下已经达到最大值。如果非生物因素,如水可用性或二氧化碳供应没有限制,碳水化合物代谢的调节特征可能会成为限制。
在光合作用过程中,光合产物以磷酸三糖的形式从叶绿体中输出,这些磷酸三糖被浓缩成己糖,最终转化为蔗糖。蔗糖通常被输出到植物的下沉器官,如生长叶片、根系和储存器官。在调节方面,两种代谢物是至关重要的。第一种是无机磷酸盐,它是从果糖二磷酸和蔗糖合成的最后一步中释放出来的,并且需要作为从质体中输出的每一个磷酸三糖的运输当量。第二种是果糖2,6-二磷酸(f -2,6- pp),它由果糖6-磷酸(F6P)产生,是果糖二磷酸酶(FBPase)活性的调节剂,催化蔗糖合成的主要调节反应。
在蔗糖在细胞质中积累的条件下,果糖6-磷酸(F6P)浓度增加,从而产生f -2,6- pp并抑制F6P的产生。这抑制了磷酸三糖从质体中提取,并在大多数植物中启动了短暂的淀粉生产。或者,蔗糖的积累可以通过输入到液泡中来避免(图15.1)。
代谢物控制反应的事实使基因修饰途径复杂化。试图通过水解焦磷酸盐来增加通量以迫使果糖-6-磷酸1-磷酸转铁酶(PFP)反应产生F6P的尝试失败了,因为在蔗糖运输的后期步骤中需要焦磷酸盐来下沉组织
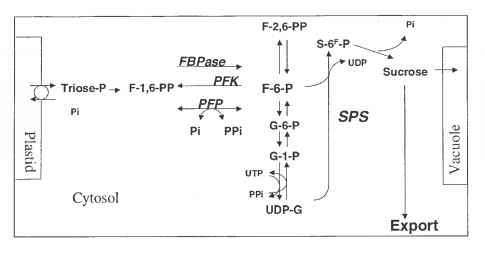
Fig.15.1。叶肉细胞(源组织)碳水化合物代谢的调节。代谢物:F-6-P:果糖-6-磷酸;f - 1, 6-PP:特性,6-bisphosphate;f - 6-PP: fructose-2 6-bisphosphate;G-1-P: glucose-1-phosphate;G-6-P: glucose-6-phosphate;P:磷酸;Pi:无机磷酸盐;PPi:焦磷酸; S: sucrose; S-6-P: sucrose-6-phosphate; Triose-P: triose phosphates; UDP: uridine diphosphate; UTP: uridine triphosphate. UDP-G: UDP-glucose. Enzymes: FBPase: fructose-bisphosphatase; PFK: phosphofructo-kinase; PFP: fructose-6-phosphate 1-phosphotransferase; SPS: sucrosephosphate-synthase.
Fig.15.1。叶肉细胞(源组织)碳水化合物代谢的调节。代谢物:F-6-P:果糖-6-磷酸;f - 1, 6-PP:特性,6-bisphosphate;f - 6-PP: fructose-2 6-bisphosphate;G-1-P: glucose-1-phosphate;G-6-P: glucose-6-phosphate;P:磷酸;Pi:无机磷酸盐;PPi:焦磷酸; S: sucrose; S-6-P: sucrose-6-phosphate; Triose-P: triose phosphates; UDP: uridine diphosphate; UTP: uridine triphosphate. UDP-G: UDP-glucose. Enzymes: FBPase: fructose-bisphosphatase; PFK: phosphofructo-kinase; PFP: fructose-6-phosphate 1-phosphotransferase; SPS: sucrosephosphate-synthase.
另一种控制蔗糖产生的方法是该途径中最后一种酶的活性,即蔗糖磷酸合酶(SPS),它通常具有低激活状态,由宿主调控。SPS的翻译后磷酸化降低了对底物UDP-G的亲和力。基因改造也受到阻碍,因为酶的过度表达不一定会增加它的活性
继续阅读:水槽容量
这篇文章有用吗?