磷虾体内持久性污染物的生物积累
在南大洋的边缘冰带是E. superba(目前占主导地位的草食动物)的苗圃,它将纳米浮游生物和硅藻产生的有机物输送给头足类动物、鱼类、海鸟、海豹和须鲸。正如在第3章中讨论的,磷虾有许多独特的特征——它的生命周期紧密匹配季节性周期在初级生产力方面,成鱼比其他远洋草食性甲壳类动物更大(可达6.2厘米长),它们具有异常高的代谢率和游泳速度,适应任何可用的食物,并在冬季食物供应低的情况下生存(Marschall 1988)。euphausiids,特别是euphausiids的一个显著特征是形成密集的聚集物,这些聚集物在面积(从几平方米到1,000平方米,有时甚至许多平方公里)、密度(从0.5到几公斤m-3)和深度(通常在水柱上方100-200米;诺克斯1994)。不同类型的聚集,如斑块、浅滩、鱼群、蜂群或超级蜂群,由不同大小和性腺发育的个体组成,它们可能是短暂的,持续数小时或数天,也可能持续数周或一生。鉴于磷虾的这些特征及其在南极远洋食物链中的中心位置,自18世纪开始海豹捕食以来,磷虾就一直受到人类的极大关注。然而,尽管进行了密集的研究,磷虾在南大洋的次生生产力及其总生物量仍然未知。根据食肉动物的年消耗量(分别为40、100、40、25和15x106吨),减少鲸鱼、海豹、乌贼、鸟类和鱼类;Knox 1994)给出了磷虾年产量的概念,以及磷虾在将金属和持久性有机污染物转移到南极远洋食物链更高层次方面的作用。
在20世纪60年代,人们对南极磷虾的捕捞可能在世界蛋白质短缺中扮演的潜在角色进行了大量的推测。大规模商业捕捞始于20世纪70年代,1982-1983年南方夏季达到高峰(528,201吨)。此后捕鱼量下降,在过去十年中稳定在每年10万吨左右(CCAMLR 2003年)。虽然磷虾可以大量捕获并加工成人类的食物,但目前全球磷虾的产量主要受到需求不足的限制(Nicol和Endo 1999)。使用磷虾作为食品的最意想不到的挫折之一是其外骨骼中的F含量非常高(高达2440 |g g-1干重量;Soevik和Braekhan 1979)。采用了将温度降低至-40°C、分离体液和煮沸甲壳类动物等程序,以减少F向冷冻磷虾肌肉组织的迁移(Knox 1994年)。尽管在开发供人类消费的南极磷虾产品方面做出了相当大的努力,但近年来,大部分收获的磷虾都被用作食品水产养殖饲料中,高F水平不是问题。
就南极磷虾的总存量和自然捕食者的需要而言,目前南极磷虾的开采水平似乎并不过分。然而,不能排除浮冰北部极限的局部种群已经被过度开发(Everson和Goss 1991年)。事实上,正如Hewitt和Low(2000)所讨论的那样,磷虾捕捞主要集中在陆地繁殖的聚居地附近磷虾捕食者.
多位作者(例如Stoeppler and Brandt 1979;山本等,1990;帕尔默·洛卡尼尼和普雷斯利1995;Barbante et al. 2000)测定了来自南大洋不同区域的E. superba样品中的微量元素浓度。尽管样品是在不同的地点和不同的季节捕获的,尽管有证据(Rainbow 1989)表明,磷藻中Cd、Zn、Fe和Mn的浓度与大小有关,并且铜含量在蜕后下降(Nicol et al. 1992),但现有数据并没有显示出相当大的变化。在大西洋东北部(Ridout et al. 1989)收集的另一种紫藻属(Meganyctiphanes norvegica)元素组成的主要差异是低锌
图45。南大洋不同区域的磷虾中微量金属平均浓度(|g g-1干wt.)与大西洋东北部和地中海中部磷虾的平均值(参考文献见正文)的比较
图45。南大洋不同区域的磷虾中微量金属平均浓度(|g g-1干wt.)与大西洋东北部和地中海中部磷虾的平均值(参考文献见正文)的比较
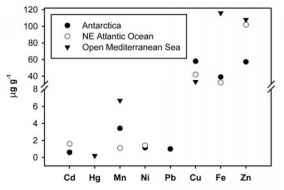
和Cd含量(图45)。地中海中部的磷虾中Fe、Hg、Mn和Zn的平均浓度(Fowler 1986)高于E. superba。后一种差异可能反映了南大洋亲石元素的生物利用度低。Barbante等人(2000)还测定了南极磷虾中Cr、Co、Sn、Se和V的平均浓度,其值(分别为0.5、0.07、0.2、7.8和1.2 |g g-1干重)与Fowler(1986)报告的在地中海收集的磷虾的浓度范围相同。
对比表13和图45的数据可以看出,E. superba在南极浮游动物中积累的Cd浓度低于其他甲壳类动物。但是,由于E. superba相对于其他甲壳类动物的生长速度较高,体型较大,这种比较只能提供一个指示,并不完全可靠。例如,Reid(2001)发现,在南乔治亚,在10月至3月期间,E. superba的大小从约42毫米增加到约54毫米,2岁的个体达到了与南极半岛3岁磷虾相同的大小。随着生物体尺寸的增大,其表面积/体重比和表面吸附金属的比例减小。此外,已知Cd主要积聚在消化腺(或脊椎动物的肝脏)和肾脏(如Bargagli et al. 1996b),成年E. superba的这些器官相对于肌肉、脂肪和性腺的比例低于其他南极甲壳类动物。因此,如果在磷虾的整个个体中测量持久性污染物的浓度,并以体重为单位来表示,即使环境或单个器官中的污染物浓度保持不变,它们也可能在个体的生长过程中不断变化。
在1975年沿南极半岛采集的磷虾样本中,Rise-brough等人(1976)测量了p,p'-DDE、p,p'-DDT和总多氯联苯含量为14、19和3 ng g-1脂质质量。滴滴涕:滴滴涕比值表明未代谢滴滴涕的比例很高。总的来说,数据为鸟类和哺乳动物特有的南极地区显示DDT及其衍生物的含量较早期有所增加
18世纪60年代至80年代初(UNEP 2002a)。然而,Bidleman et al.(1993)报告称,在1990年的样品中,新鲜DDT的大量投入仍然很明显。
Sen Gupta等人(1996)在1987年12月至1988年1月在大约70°S和12°e收集的E. superba样品中测量到t-六氯环己烷(a+g异构体=0.154 ng g-1干质量)的平均浓度高于总多氯联苯(0.152 ng g-1干质量)和总滴滴涕(p,p'-DDT+o,p'-DDT+p,p'-DDE+o,p'-DDE=0.037 ng g-1干质量)。滴滴涕、p,p'-DDT和p,p'-DDE的主要异构体和代谢物在磷虾体内的比例几乎相等。主要的PCB同源物是PCB-138和PCB-136。Corsolini等人(2002)于2000年1月测量了罗斯海(71°20’s -72°331’s /170°22’e -178°04’e)磷虾样品中p、p’-DDE和HCB的平均浓度(分别为0.86±0.98和0.37±0.17 ng g-1 wet wt.)。这些样品中的PCB平均浓度要高得多(167±85 ng g-1湿wt.),同族特异性PCB图谱显示低氯异构体普遍存在(四氯联苯占大部分残留)。这种模式不同于通常在低纬度和中纬度生物中检测到的模式,这可能是由于高纬度地区的全球分馏。来自罗斯海的生物的异构体模式与Kanechlor相似,Kanechlor是一种技术上主要用于日本和其他东亚国家的混合物,大致位于罗斯海的经度(Corsolini et al. 2002)。雷竞技手机版app以脂质重量为基础,同一磷虾样品中总多氯二苯并对二恶英(pcdd)和二苯并对呋喃(DFs)的浓度为27 pg g-1,非和单邻位取代多氯联苯(二恶英类多氯联苯)的浓度为0.9 ng g-1。
1994年11月从罗斯海采集的E. superba样品的人工放射性核素含量非常低(<0.11 Bq kg-1干重的137Cs),而90Sr、239Pu、238Pu和241Am则检测不到(Nonnis Marzano et al. 2000)。
为了评估几种生物标记物(酯酶、混合功能氧化酶、卟啉)的潜在用途,Minutoli等人(2002)分析了地中海和罗斯海整个浮游甲壳动物匀浆中的乙酰胆碱酯酶(AChE)活性。在磷藻类中,来自地中海的褐家蝇AChE活性(13.3 |mol min-1 g-1)高于E. crystorophia和E. superba(分别为4.5和1.7 |mol min-1 g-1)。这种差异归因于低的基础代谢活性南极生物而E. superba相对于E. crystorophias的活性较低可能是由于它们的尺寸不同(分别为62和34 mm)。事实上,已知AChE活性与生物体尺寸成反比(Fossi et al. 1996)。
继续阅读:全球环境挑战和减少对南极洲的不利影响
这篇文章有用吗?