二氧化碳
我们将首先考虑二氧化碳的程度可用性限制了总体增长速度。因为二氧化碳是一个使用酶的底物(系统),然后我们可以合理的假设光合速率在任何给定的光强度将随二氧化碳浓度大约依照知名Michaelis-Menten酶动力学方程
公里+ s v是酶反应的速度在底物浓度,v是最大的饱和率渠道底物浓度和公里的离解常数es复杂,但也相当于出一半的底物浓度的最大速率。Re-expressing方程在光合作用方面得到p = _Pm (C(°2) __(2) 11日
公里)+{二氧化碳(CO2) 1”2
图11.2显示了一个理想化的速度曲线与酶反应底物浓度,按照MichaelisMenten方程。观察曲线的光合速率与二氧化碳
水生植物的浓度大约这种类型的,所以得到的Km值曲线和原位二氧化碳浓度进行比较,我们可以评估在多大程度上二氧化碳的可用性限制光合作用在自然水域。
在孤立的状态,二磷酸核酮糖羧化酶从水生植物对二氧化碳30公里价值mM.1489 70公里为植物(CO2)值,然而,可以更高,因为二氧化碳的扩散阻力进入到植物,或降低由于积极吸收二氧化碳。值公里(CO2)决定对浮游植物和大型植物物种,在pH值足够低(pH < 6)以确保基本上所有的无机碳(Ci)存在二氧化碳(或其水合形式,H2CO3),表11.1列出了:他们从4到185 mM。淡水平衡与大气中二氧化碳(^ 0.035卷%)15°C包含溶解二氧化碳浓度约为14毫米。计算海水26°C显示免费的二氧化碳浓度约12 mM.136因此我们可能期望它通常是在自然水域的水生植物光合作用是欠饱和对二氧化碳浓度,而且应该应对二氧化碳浓度的增加与photo-synthetic率的增加。这个不需要只适用于饱和光强:假设光合速率取决于二氧化碳浓度按照Michaelis-Menten关系意味着,即使在低光强度的增加二氧化碳浓度超过12到14毫米的典型起始值会导致光合速率的增加。这意味着在subsaturating光强度,光合作用植物在水体可能同时受限于可用性的光和二氧化碳。水生苔藓,Fontinalis antipyretica,水生高等植物,Cabomba虎耳,都显示出显著增加光合速率随着二氧化碳浓度在10到25毫米范围低,subsaturating,以及在高,光强度。1235年523年,图11.3显示了Cabomba史密斯(1938)的数据。然而,这些例子是有些不地道,因为大多数水生高等植物和藻类,在特定水域的自然发生,并不显示应对二氧化碳供应的增加。原因是水生植物不仅可以利用其他无机碳的来源,但他们中的大多数也有能力执行Ci的主动运输,这是通常被称为carbon-concentrating机制,或CCM。这个话题由法了,乌鸦(2007),由卡普兰和莱因霍尔德(1999),并在《第四届国际研讨会上无机碳利用水生光合生物,2001(功能性植物生物学,2002卷。29)。
表11.1明显half-rate常量(公里)CO2for光合作用在某些浮游植物和大型植物物种。
植物物种
淡水浮游植物
绿藻门
盘星藻boryanum 40
鼓藻170年葡萄孢属
衣藻reinhardtii 29
栅藻obliquus 38裸藻门植物
眼虫属股薄肌25蓝藻门
项圈藻cylindrica 60
丝囊藻属flos-aquae 105
Coccochloris peniocystis 121
织线藻属boryanum 100
淡水大型植物
Nitella flexilis 100
Eurhynchium rusciforme 80
Fontinalis antipyretica 170
伊乐藻属植物黄花30
Potamogeton管20
软水草verticillata 170
Myriophyllum spicatum 150
Ceratophyllum demersum 165
海洋浮游植物
绿藻门
Stichococcus bacillaris 4 Heterokontophyta
Cylindrotheca梭杆菌属(硅藻)36
Olisthodiscus危害(raphidophyte) 59红藻门
紫球藻属cruentum 22蓝藻门
聚球藻属sp 240海洋大型植物
石莼sp。30
石莼以185
384年19日19日384年
19日384 384 384
19日19日19日19日19 1394 1394 1394
384 384
86 321
海水和大多数内陆水域包含更多的无机碳的形式碳酸氢盐离子,HCO3 ~比以二氧化碳的形式。不同形式的无机碳在根据互换
水+ H +二氧化碳^ H2CO3 ^ + HCOiT ^ 2 H + + CO32 ~
-
.")
- 图11.3光合速率作为二氧化碳浓度的函数在不同的光强度的水生高等植物Cabomba虎(Rabinowitch之后,1951年,基于数据的史密斯,1938)。
.")
水的pH值越高,越这平衡转移到正确的HCO3 ~构成超过总数的50%从pH值6.2至9.3,以及pH值6.7和8.8之间的超过80%。鉴于二氧化碳倾向,正如我们所见,出席非最优浓度,它显然是一个优势在许多水域水生植物能够使用碳酸氢钠作为光合作用的碳源,事实上很多,但不是全部,物种。使用的无机碳物种羧化酶酶都是二氧化碳。植物可以利用碳酸氢盐,HCO3 -离子输送到细胞,产生二氧化碳的逆转上述前两个互变现象的反应。可逆的脱水H2CO3给二氧化碳是由碳酸酐酶催化,酶发生在叶绿体,但也可以出现在细胞质内和胞质膜的外表面。然后用于光合作用释放二氧化碳。在外部酶的情况下,释放二氧化碳扩散通过细胞质膜进入细胞。HCO3 ~与H +离子的结合给H2CO3导致相应的氢氧根离子的积累(从水!H + +哦),从细胞分泌平衡HCO3的吸收。
所以海洋生态系统而言,光合利用碳酸氢被发现在大多数的海草,绿色,棕色和红色——已经被研究过,但并不普遍。Maberly(1990)发现的35种海洋大型六个物种-所有rhodophytes无法使用HCO3 -和五个发生在相对低光照条件下的栖息地,树冠下的大褐藻类。他建议大多数物种生长在深度光HCO3低将无法使用。红色macroalga陨石球粒管,虽然它不直接占用碳酸氢盐,然而获得访问这个丰富的无机碳池通过脱水HCO3 ~二氧化碳与外部碳酸酐酶。1254年、1255年的29 macroalgal物种(11 13绿色,5红、棕色)研究了拉尔森和Axelsson(1999)所有除了一个(Palmaria palmata,红色)利用HCO3 ~通过外部碳酸酐酶。
除了能够利用HCO3 ~通过外部碳酸anhy-drase,一些藻类也有能力积极运输离子进入细胞。两个常见的和成功的绿色藻类物种,Cladophora glomerata和浒苔intestinalis,都有碳酸氢盐转运蛋白。229771一些,但不是全部,绿色藻类物种有碳酸氢诱导运输车,运营阴离子交换:770229这种能力的发展在高博士是缺席的布朗和红藻物种测试。褐藻,Hizikia fusiforme,具有外部碳酸酐酶,但不进行主动运输的HCO3 ~。1506 Choo et al。(2002)发现,除了外部碳酸酐酶和离子交换类型碳酸氢盐,Cladophora glomerata也可以形成一个运输机使用质子泵atp酶的类型。质子是electrogenically抽出单元创建一个活跃的质子梯度,然后导致进口运输的H +和HCO3。质子泵的形成是引起的碳排放限制。额外的机制,利用质子梯度的本地化结合H +和HCO3 ~给H2CO3的二氧化碳解放通过外部碳酸酐酶。这似乎存在于charophyte,性格最纯粹,393年,1088年和褐藻、海带saccharina。41所产生的二氧化碳从而存在局部高浓度,导致跨细胞膜向内扩散。在催化质子分泌发生在专门的酸性乐队。这些都伴随着不同碱性bands1114碳酸钙硬壳发生:这些可能对应地点哦分泌H +(或吸收)发生雷竞技csgo保护碱度。85 Even in those macrophytes, such as Ulva, which can utilize bicarbonate it seems that photosynthesis in bright light can in some cases be limited by the level of inorganic carbon in sea water.794a
碳酸氢盐利用率是发现在一些,87847,1092,但显然不是全部,5海草。外部,膜结合碳酸酐酶在这些海洋被子植物中很常见。85年这有助于吸收无机碳不构成CCM。似乎很猛,那些可以进行主动运输的海草的碳酸氢盐通过一个质子泵创建一个H +梯度。这种梯度可以用来带来碳酸氢盐运输两种方式。可以有内在的协同转运的H +和HCO3,或可能有局部结合H +和HCO3 ~给H2CO3的二氧化碳通过外部碳酸酐酶,解放了393年,1088年,85年后跟跨细胞膜向内扩散作为性格的描述,以上。和性格一样,似乎是在空间上分开的酸和碱性区域。85这个特殊的CCM机制不是万能海草:热带物种研究的八个Uku et al .(2005),这是存在于6 -在两个缺席。目前,利用这种机制,卡尔和Axelsson(2008)发现,ATP的主要部分用于生成酸区是来自线粒体呼吸。海草不似乎拥有一个活跃的阴离子交换类型的传输机制。
同位素不平衡实验的基础上,进行浮游植物从东部亚热带和赤道太平洋Tortell和龙葵(2002)认为HCO3无机碳的主要形式是被所有的原位人口抽样。所有真核浮游植物物种包含内部碳酸酐酶:外部碳酸酐酶的存在是常见的但不普遍。海洋浮游植物中HCO3 ~利用已被证明在coccolithophorid Coccolithus huxleyi; 1028年的硅藻,haptophyte eustigmatophyte物种;256年,在单细胞红藻类紫球藻属cruentum; 255年绿色微生物Stichococ-cus bacillaris具有高度的亲和力二氧化碳(表11.1)和低亲和力碳酸氢盐,966和一些常见的海洋硅藻似乎无法使用碳酸氢盐。296年、1127年Dason et al。(2004)发现,两个海洋甲藻、Amphidinium carterae和Heterocapsa大洋洲,不能占用碳酸氢盐,但似乎有一个主动转运系统二氧化碳本身。似乎也这样两个海洋绿色植物物种,Nannochloris atomus和n maculata。598共生甲藻(Symbiodinium sp黄藻)在巨大的蛤蚌CCM,和优先运输CO2.792 Burkhardt et al。(2001)目前的证据表明两种海洋硅藻,这种weissflogii和Phaeodactylum tricornutum,拥有活跃的HCO3和二氧化碳的运输系统。rhodophyte,紫球藻属cruentum,还积极传输形式的词,598年淡水eustigmatophyte, Eustigmatos vischeri, 597和某些海洋prymnesiophyte物种。108596抑制剂的研究表明,一种阴离子交换的HCO3 ~交通系统发生在某些真核海洋浮游植物物种,而不是其他人的。992 Maberly et al。(2009)测试物种从12金藻家庭占用无机碳的能力。结果表明,金藻集团没有CCM,此外没有能力利用碳酸氢作为替代无机碳的来源。
海洋蓝细菌,如聚球藻属、主动运输HCOr和二氧化碳转化为细胞。细胞内的55岁,1089年,56 Ci HCO3主要存在。碳酸氢盐离子扩散到carboxysome,半晶状的结构,主要包括二磷酸核酮糖羧化酶,还包含碳酸酐酶。后者酶保持高本地化水平的二氧化碳从HCO3通过不断生成,这是足够的饱和羧化酶,竞争性抑制Rubisco.384的加氧酶活动
在新鲜的水域,根据哈钦森(1975)评论,大多数水生高等植物可以利用碳酸氢进行光合作用。Maberly和马德森(2002),在他们的淡水被子植物的碳吸收机制,报告使用HCO3 ~大约一半的水下被子植物物种的存在,测试,和更常见的高碱性湖泊。物种,不能利用碳酸氢通常发生在软水域中碳酸氢盐浓度很低。水生苔藓主要利用HCO3似乎缺乏能力。大多数charophyte物种(从软水域以外)可以利用HCO3 ~ Cladophora等可以底栖生物丝状藻类。淡水浮游植物,利用HCO3 ~进行光合作用的能力差别很大。小球藻emersonii,栅藻quadricauda莱恩衣藻(绿色性状进行),角藻hirundinella(腰鞭毛虫)Fragilaria crotonensis(硅藻),微胞藻属绿脓杆菌和项圈藻cylindrica(蓝)都可以有效地使用HCO3但小球藻pyrenoidosa(绿色),Asterionella福尔摩沙和株italica cannot19,839(硅藻),1109年,1338年,1377年某些其他淡水小球藻的物种,和淡水硅藻,舟状窝pelliculosa,开展主动运输的HCO3 -和CO2.256
尽管碳酸氢能力利用的广泛出现在水生植物王国,它通常是事实,可能除了浮游植物物种适应碱性水,免费的二氧化碳,如果是首选,即更有效地使用,碳源。例如,淡水大型植物Myriophyllum spicatum,可以使用碳酸氢盐,给出了一个更高的光合速率与自由二氧化碳的最佳浓度比以HCO3 ~最佳浓度。1297。博得纳(1994)发现水池草,Potamogeton•,使用二氧化碳,但无法使用HCO3:毛茛属植物fluitans优先使用二氧化碳,但能够切换到HCO3 ~ 9.0及以上的pH值,当二氧化碳浓度接近于零。Steemann尼尔森(1975)表明,有效使用越少的原因之一HCO3 ~是能源必须用于主动运输到工厂,而二氧化碳在自由扩散。艾伦和斯宾塞(1981)的数据淡水大型植物在表11.1中显示,在所有情况下,包括已知的碳酸氢盐用户伊乐藻属植物黄花和Potamogeton管,明显公里HCO3 ~ 50 - 100倍(显示更低的亲和力)高于二氧化碳的公里。此外,给光合速率与一定浓度的二氧化碳(pH值5.5),HCO3 ~ (pH值8.8)在52到132倍浓度是必要的。的蓝绿色藻类项圈藻cylindrica,另一方面,通常发生在碱性富营养化水二氧化碳的Km值和HCO3 ~差不多,和二氧化碳的浓度和HCO3 ~产生一定的光合速率也大致相同。学业的基础上,艾伦和斯宾塞认为,尽管大多数淡水植物利用碳酸氢的能力,事实上他们并没有获得他们的大部分碳HCO3 ~直到水的pH值超过9.0,并在这些高pH值光合率大大降低。艾伦和斯宾塞认为,自然利率淡水大型植物和一些浮游藻类光合作用的功能主要是二氧化碳(而不是HCO3)浓度。然而,这一结论并不适用于大型植物之间的更有效的碳酸氢盐用户,比如Myriophyllum spicatum。亚当斯et al。(1978)发现,在一系列的,而碱性(pH值主要是7.5 - -8.8)意大利湖泊,所有与磷,供应充足的光合作用速率m . spicatum多样
总经销(}iw«d inoraaoic碳水化合物«(mM |□。(J 1我詹。3 D乔40
0 5 q 100杠杆收购2 (K
总di [Hlv«我无机carimn(国家行业集团公司有限公司]equig ^ Eenn
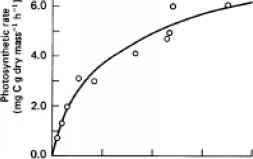
图11.4光合速率变化Myriophyllum spicatum总溶解无机碳浓度在一系列的意大利湖泊(数据绘制的亚当斯et al ., 1978)。光强度在大多数情况下都达到或接近饱和。MichaelisMenten方程使用非线性回归,亚当斯等人计算公里(half-saturation常数)值的1.06毫米总溶解无机碳(46.5毫克二氧化碳等价物l1)和点值为7.24毫克C g_l h_1干燥质量。
0 5 q 100杠杆收购2 (K
总di [Hlv«我无机carimn(国家行业集团公司有限公司]equig ^ Eenn
图11.4光合速率变化Myriophyllum spicatum总溶解无机碳浓度在一系列的意大利湖泊(数据绘制的亚当斯et al ., 1978)。光强度在大多数情况下都达到或接近饱和。MichaelisMenten方程使用非线性回归,亚当斯等人计算公里(half-saturation常数)值的1.06毫米总溶解无机碳(46.5毫克二氧化碳等价物l1)和点值为7.24毫克C g_l h_1干燥质量。
总溶解无机碳浓度按照Michaelis-Menten类型的关系(图11.4)。从计算量的不同形式的无机碳,看来光合速率主要是相关的浓度HCO3 ~而不是二氧化碳的浓度。
琼斯(2005)指出,一个活跃的交通系统的形成和操作HCO3 ~必须对植物的能源成本。与伊乐藻属植物实验nuttallii,生长在水中含有的碳酸氢盐、和充溢正常的空气或空气耗尽的二氧化碳(这将有利于转向HCO3 ~使用),他发现后21天bicarbonate-using植物光合作用在^率高58%,但只取得了相同的增长主要植物利用二氧化碳,这表明一些光合成的捕获光能被用于构建、维护和运行碳酸氢盐利用系统。琼斯估计这将对应领域的辐照度~ 80 m ~ 2 s_1更易与光子,低于HCO3 ~使用不会期望在这个物种。因为这是水下的辐照度通常有经验的大型植物在内陆水域,HCO3 ~使用可能会局限于地区环境光超过必要的阈值。
项圈藻等藻类cylindrica,光合作用最佳pH值在9.0的区域,可以非常有效地使用碳酸氢盐。对于这些浮游物种和更有效的碳酸氢盐用户在大型植物,自然的光合作用(如碳供应而言)可以被视为一个函数的HCO3 ~ +二氧化碳的浓度。项圈藻等蓝藻的能力,微胞藻属和螺旋藻继续在水pH值高、光合作用和有效零二氧化碳浓度可以自由的一个主要因素频繁的主导地位湖泊富营养化在夏末和不变的高碱性水,例如非洲的统治碱湖。一些蓝藻形成表面大量繁殖的能力给他们一个额外优势:通过将一个密集的生物质接近水面,它们能够拦截大部分传入的大气中的二氧化碳,以及入射光.607
Rattray et al。(1991)发现,两根淡水植物,Lagarosiphon主要和Myriophyllum triphyllum,增长了两倍,photosynthe-sized贫瘠陶波湖的水(新西兰),部分归功于富磷氮水的富营养的罗托鲁瓦湖。Rattray等人将此归因于两方面的更高层次的二氧化碳被发现出现在贫瘠,比富营养水。软水贫营养湖泊常常为底栖大型植物的存在与isoetid增长形式:较短的小植物茎和叶子的花结。603 Isoetids已经解决了这个问题了这些糟糕的无机碳浓度低矿化水从沉积物中提取的二氧化碳,形成从腐烂的有机物质,通过他们的根源。142847、1雷竞技csgo111、846这二氧化碳扩散从根到叶子通过纵向充气通道,并在某些情况下提供了几乎所有的光合成固定碳。紧急大型植物,莎草纸莎草纸,也有大型细胞间空气腔(通气组织)延伸其沉积物中植物的根状茎,和能够光合成捕获二氧化碳从rhizomes.808上来
12到14毫米的自由二氧化碳浓度称为早期仅适用于水与大气平衡。通常这样的浓度可能被发现在海洋的表层,或非生产性内陆水域,冬天大多数内陆水域。然而,任何水体生物生产力不太可能是在平衡大气二氧化碳而言。随时免费的二氧化碳浓度可能不同于平衡值,并将随时间和深度依照地方和消费的过程有多快(光合作用)和生产(呼吸、分解)。模型的基础上计算利率的二氧化碳扩散的细胞,和实验室测量的增长率随着二氧化碳浓度的函数,Riebesell et al。(1993)认为在海洋浮游植物水华期间,当二氧化碳浓度在表层大幅下跌,海洋硅藻的生长速率可以通过二氧化碳供应有限的细胞表面。Vadstrup和马德森在贫营养湖低碱(1995)发现Hampen(丹麦),免费二氧化碳的浓度大约五倍大气平衡浓度在初夏,但拒绝夏天年底几乎为零。
免费的二氧化碳可以更受pH值的变化引起的光合作用。安装(1976,1979)提出了数据的免费的二氧化碳浓度的变化英语湖富营养化,Esthwaite水,从4月到1971年7月。4月19日,之前热分层现象,二氧化碳浓度是一样的从表面到10米深度。5×3浮游植物数量开始增加,并有一些表层的变暖,二氧化碳浓度下降了大约33%在上4米但大大增加了6 - 10米。在7月,热分层现象成为建立,浮游植物种群在温跃层中二氧化碳浓度大幅增加和自由变温层减少几乎为零,而上升到更高的价值比以前在6到10米层温跃层以下。无机碳的数量被光合作用代表了大约50%的总无机碳的表层。~ 1000倍的其余部分免费的二氧化碳减少是由于pH值的上升从^ 7.0到9.0以上,造成人口密集的浮游植物的光合作用的活动。相似的变化可能发生在任何生产水体。浮游植物的组件之一,硅藻Asterionella福尔摩沙(不是碳酸氢盐的用户),安装(1979)7月12日计算,其光合速率几乎为零的表层由于缺乏二氧化碳,它上升2和3 m之间由于二氧化碳的增加伴随着相当高辐照度值,并拒绝再次低于这个深度,因为辐照度的下降。因此随着深度增加,其光合作用被二氧化碳供应有限的第一,然后同时通过二氧化碳和光线,主要是由光的可用性。这些变化与深度并不适用于总浮游植物的光合作用,然而,因为当pH值上升和二氧化碳浓度下降,Asterionella主要是碳酸氢取代用户角藻和微胞藻属。
硅藻细胞的下沉率增加当二氧化碳吸收有限:二氧化碳相关损耗夏季分层可能导致硅藻的下沉率增加,这可能导致硅藻经常观察到的缺乏生产力summer.622湖泊
立即任何水生植物表面的旁边,有一个没有被搅动的层水,二氧化碳和/或HCO3 -离子必须分散在之前进入细胞,可用于光合作用。这一层的厚度减少,如果植物受到湍流或快速搅拌但层从未完全消失。搅拌好情况下,没有被搅动的层可能只有5毫米厚约一个小细胞,如小球藻但30至150毫米厚的表面等大型植物的性格:1109年,1236年还是缓慢流动的水没有被搅动的层往往会远远超过150毫米厚。乌鸦(1970),和史密斯和沃克(1980),有了这个话题,得出扩散的二氧化碳(大概HCO3)在这个没有被搅动的层可以是一个重要的病原反应一步水生植物的光合作用。河植物光合速率的增加与增加在自然水域流速度其实已经证明了西湖(1967)。惠勒(1980 b)发现,叶片的光合速率的海带,Macrocystis pyrifera,在饱和光,增加约4倍时,当前的速度从0增加到5厘米s - 1:在低光,然而,这一增长仅为50%。任何形态适应增加将有助于克服扩散问题的表面体积比和许多水生大型植物物种发展的高度解剖叶子已经达到了这个要求。603年切割叶形式越多,越少的刺激增加光合作用所带来的动荡。451是否存在一层没有被搅动的每个细胞都有任何重大限制浮游植物光合作用的影响仍不确定。
虽然淹没淡水大型植物的光合作用速率会增加反应增加当前的速度大约1 - 0.5厘米,可以有抑制效应对水速度超出^ 0.5 - 3厘米s - 1.845 - 14,991使用放射自显影法co2-exposed伊乐藻属植物的叶子黄花,尼尔森et al .(2006)发现,在高流速的二氧化碳吸收大约是双重的更高的叶片边缘与中脉附近的部分,而在低流速相对较高的二氧化碳吸收的面积扩大从叶子边缘向中脉和基底的部分。他们建议缓慢的水会在叶技巧和进入螺环树叶间的缝隙,实现二氧化碳的基底部分阀杆附近的树叶。在高速度的水不进入空白,而是跳过,有效提高表面的水平。这发生的程度很可能是植物形态学的函数。
在大多数陆生高等植物物种,那些缺乏C4途径,由氧光合作用显著抑制正常大气水平(21%),主要是由于O2和CO2之间的直接竞争的二磷酸核酮糖羧化酶的活性部位。二磷酸核酮糖羧化酶的oxyge-nase反应的氧与二磷酸核酮糖反应产生phosphoglycollate和磷酸甘油酸酯,这是第一步被称为光呼吸代谢途径。由水生高等植物和藻类光合作用,相比之下,在大多数情况下显示了氧气抑制相对较少。这似乎是由于水生植物的能力增加,由一个或其他一系列机制,二氧化碳的浓度接近的二磷酸核酮糖羧化酶分子在细胞内,从而增加二氧化碳/ O2的竞争比。正如我们所看到的,证据的主动运输二氧化碳,和HCO3 -在某些情况下,内部实现大大高于外部浓度。
以及“生物物理”战略有生化问题的解决方案提高内部的二氧化碳浓度。水下水生植物尚未发现的展品C4光合作用发生在陆地C4物种,即固定的二氧化碳转化为C4植物叶肉细胞酸苹果酸等,其次是运输这种酸专门的维管束鞘细胞CO2释放然后断代为正常光合碳水化合物的碳减排(PCR)循环。然而,可能被视为一个结构的微缩版C4途径发现萨吴奇和Bowes(1983)在淡水大型植物软水草verticillata (Hydrocharita-ceae),整个生化序列似乎发生在相同的细胞。也有证据表明这种C4光合作用在某些水鳖科的其他成员,包括软水草verti-cillata852和女顾问densa。212在这两种情况下,形成C4循环的酶是诱导植物生长条件下(高温、高亮度、长光周期),导致限制二氧化碳水平在水里。C4的审查机制在水生被子植物,Bowes et al。(2002)将软水草称为兼性C4转变从C3, C4在低[CO2]。
似乎大部分的初始固定二氧化碳是由磷酸烯醇丙酮酸(PEP)羧化酶在细胞质中。oxaloa-cetic酸生产降低苹果酸。这是运入叶绿体在脱羧的辅酶ii苹果酸脱氢酶,二氧化碳是固定的PCR循环的碳水化合物。149这种类型的证据,C4途径被发现在一个海草物种Cymodocea nodosa,但不是在十人。88也有证据C4途径,但使用PEP carboxykinase羟酶,在海洋绿色宏观藻类Udotea flabellum.1118雷竞技csgo
浮游植物,Reinfelder et al。(2000)证明短期C4化合物的标签,和转让碳糖,在海洋硅藻这种weissflogii,解释为C4光合作用的证据在这个物种。Reinfelder et al。(2004)发现,特定的PEP羧化酶抑制剂引起细胞的光合作用下降超过90% t weissflogii适应低[CO2],但影响很小C3-using海洋绿色植物,衣藻sp。类似的抑制实验的基础上,作者和龙葵(2008)建议C4-based CO2-concentrating机制raybet雷竞技最新通常分布在硅藻,表明他们拥有C4-based CCM可能导致的成功进化phyto-plankton门,尤其是在较低的二氧化碳浓度,描述今天的地表水。
另一个生化CO2-concentrating机制,景天酸代谢(CAM),发生在一些,但不是全部,isoetid的大型植物种类的贫瘠,软水、湖泊。回顾了14268雷竞技csgo1年、1111年的光合生理马德森et al。(2002)。这些水生CAM植物有玫瑰的短硬叶子和大根可以超过50%的植物生物量。通过他们的广泛的根系吸收二氧化碳的沉积物,在高度集中的地方。运输的二氧化碳从根到叶子发生扩散通过系统内部气体的空间(缺损)。大部分的无机碳用于光合作用isoetids来自沉积物二氧化碳,它都是在夜间和白天一样。拥有凸轮使这些植物修复二氧化碳在夜间通过PEP羧化酶、苹果酸等有机酸的形式积累在液泡。白天,当光线可用,二氧化碳是解放C4的脱酸,和固定碳水化合物由PCR循环。
继续阅读:间接因素
这篇文章有用吗?
读者的问题
-
波西亚2个月前
- 回复
-
红桉2个月前
- 回复
-
苏菲2个月前
- 回复
-
蛋白石2个月前
- 回复
-
安德烈亚斯1年前
- 回复
-
菲利普1年前
- 回复
-
PUPETTA1年前
- 回复
-
杰西卡·李1年前
- 回复
-
克里斯多夫1年前
- 回复