叶绿体运动
光收集的速率叶绿体这不仅取决于它们的色素补体,还取决于它们在细胞内的位置和方向。在一些水生植物中,这可能随着光照强度的变化而变化。在高等植物中,一般的行为模式是,在低强度的光下,叶绿体移动到细胞中的某个位置,使光吸收是最大化的,它们向外扩散,与这些细胞壁相邻并平行入射光。在高强度光下(光合作用饱和),叶绿体移动到一个位置,使光吸收最小化,也许是为了减少光抑制:它们远离直接暴露在光线下的细胞壁,并与阴影最严重的细胞壁(通常是侧壁)对齐并平行,这样它们不仅是面朝光的,而不是正对着光的,而且有些细胞壁被其他细胞壁遮蔽。以生长在苏格兰湖中的水生被子植物Potamogeton crispus为例,从0.25米深处采集的叶子的叶绿体分布在细胞的侧壁周围,而从2.5米深处采集的叶子的叶绿体分布在面向光线的表面在高强度光下减少光收集的另一种方法是叶绿体在细胞核周围聚集成簇:这在生长在地中海0.5米深处的海草Halophila tolacea的叶子中观察到。323 Sharon和Beer(2008)比较了生长在强光(中午450 mmol光子m~2s_1)和弱光(中午150 mmol光子m~2s_1)下的tolacea植物。日出前06:00 h,叶绿体均匀分布在叶细胞的细胞质内。在强光植株中,叶绿体在日出时开始迁移,并在08:00 h观察到开始结块。正午开始完全结块,并持续整个下午。下午晚些时候,叶绿体开始在细胞质中扩散,并在日落后几小时达到完全扩散。在光照较弱的植物中,白天没有叶绿体结块。在强光植株中,伴随叶绿体结块的是叶片吸收率的下降,从清晨的0.56下降到中午的0.34,而叶绿素含量没有相应的变化。在亚喀巴湾(红海)的托叶苋植物中,Schwarz和Hellblom(2002)发现,在7米深的植物中,叶绿体结块使叶片吸收率降低到55%,而在30米深的植物中,这一比例为85%。
的光谱高等植物的叶绿体运动在蓝色区域约450nm处有一个峰:光感受器可能是一种黄素或类胡萝卜素其机制可能与细胞质流动有关:叶绿体似乎与细胞质一起移动,而不是穿过细胞质。
Nultsch和Pfau(1979)研究了大量沿海和亚沿海海洋大型藻类的叶绿体运动。在大多数棕藻物种中,叶绿体在低辐照度时移动到面向光的细胞壁,在高辐照度时移动到与光方向平行的侧壁(图12.19):从一个位置到另一个位置的变化需要1 - 2小时。在所研究的绿色或红色藻类物种中,没有观察到明显的光诱导的叶绿体运动。Horstmann(1983)在热带浅水礁区生长的虹吸状绿藻(corerpa racemosa)中观察到,在明亮的阳光下,叶绿体从叶片缩回到匍匐茎中。
-
, Marine Biology, 51, 77-82). Arrangement of the chloroplasts in (a) low-intensity (1000 lux) and (b) high-intensity (10 000 lux) light. Magnification ~4000 x.")
- 图12.19光诱导褐藻saccharina中叶绿体的运动(获得Nultsch和Pfau(1979)的许可,海洋生物学,51,77 -82)。叶绿体在(a)低强度(1000勒克斯)和(b)高强度(10 000勒克斯)光中的排列。放大倍数~4000倍。
, Marine Biology, 51, 77-82). Arrangement of the chloroplasts in (a) low-intensity (1000 lux) and (b) high-intensity (10 000 lux) light. Magnification ~4000 x.")
在丝状绿藻moougeotia和Mesotaenium中,每个细胞中都有一个位于中心的单一平坦的矩形叶绿体,叶绿体响应光状态变化的特征运动是在其纵向轴上转动,而不是绕着壁移动。它可以正面转向中光,侧面转向强光。Haupt(1973)和同事已经证明控制这种运动的感光细胞是光敏色素。在另一种丝状共胞藻类,无柄囊藻(叶藻科),如果丝在某一点被低强度蓝光照射,通常由流动细胞质携带的叶绿体和其他细胞器就会聚集在细胞被照射的部分这对光合作用的影响尚不清楚。
以漂浮的浮游藻类为例,它们随光线的方向是随机的,如果细胞中只有一个或几个叶绿体,那么仅仅移动到细胞的一个部分或另一个部分不太可能对光的收集速度产生很大的影响。在某些情况下,叶绿体可以降低其毒性吸收截面在强光下通过收缩:这已在甲藻1329和硅藻中观察到。537,686在海洋中心硅藻Lauderia borealis中,每个细胞约有50个叶绿体,Kiefer(1973)观察到,在强光照射(244 Wm2)的前2分钟,叶绿体的大小收缩(图12.20a)。在接下来的30 - 60min中,叶绿体在弱光下均匀分布在周围,并移动到细胞的瓣膜末端,形成两个大小相等的聚集体(图12.20b)。叶绿体的大小和位置的变化伴随着悬液在440 nm处的吸光度下降了约40%(可能低估了,因为散射总是会产生一些虚假的吸光度):因此,通过收缩和聚集的结合,这种硅藻能够大幅降低其在强光下的能量收集率。
Stephens(1995)研究了光诱导的叶绿体迁移和相关的光谱变化,在甲藻中,月牙藻是一种丰富的物种,在海洋的热带和亚热带地区,它有巨大的月牙形细胞。细胞在12小时的光照和12小时的黑暗循环下生长。在黑暗周期中,叶绿体位于细胞的两个远端部分(新月形的“角”)的细胞质链之间,而不在细胞的颗粒中心区域。当细胞从黑暗中取出并置于显微镜的光束下时,叶绿体开始移动,2至3分钟后聚集在细胞的中心区域。吸收光谱的
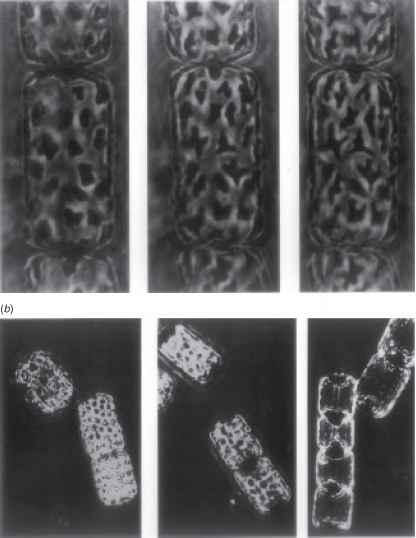
图12.20海洋硅藻Lauderia borealis高光强诱导的叶绿体收缩和聚集(经允许,来自Kiefer (1973), marine Biology, 23,39 -46)。在每一种情况下,一系列显微图显示了从左到右不断增加的光诱导变化。(a)叶绿体收缩。(b)叶绿体移动到细胞的瓣膜末端,然后聚集。
图12.20海洋硅藻Lauderia borealis高光强诱导的叶绿体收缩和聚集(经允许,来自Kiefer (1973), marine Biology, 23,39 -46)。在每一种情况下,一系列显微图显示了从左到右不断增加的光诱导变化。(a)叶绿体收缩。(b)叶绿体移动到细胞的瓣膜末端,然后聚集。

120 144
图12.21加拿大圣劳伦斯河口浮游植物光合能力(Pm)在7天内的昼夜节律(经Demers和Legendre(1981)许可,海洋生物学,66,243 -50)。
120 144
图12.21加拿大圣劳伦斯河口浮游植物光合能力(Pm)在7天内的昼夜节律(经Demers和Legendre(1981)许可,海洋生物学,66,243 -50)。
细胞中心聚集的叶绿体比细胞远端区域的叶绿体表现出更明显的包裹效应,即峰的扁平化。在这种情况下,叶绿体运动也有降低强光下能量收集率的作用。
继续阅读:的microphytobenthos
这篇文章有用吗?
读者的问题
-
feaven2个月前
- 回复
-
玛丽亚2个月前
- 回复