色适应在真核藻类
红藻表明色素成分的变化,以应对变化的光谱质量的光场他们成长。色素的方向变化,然而,除了它的量化程度,似乎取决于光的强度。布罗迪和爱默生(1959)确定的比率藻红蛋白在单细胞红藻类叶绿素紫球藻属cruentum生长在绿灯(546 nm -主要是由藻红蛋白吸收)或蓝色(436 nm主要是由叶绿素吸收),在低(Wm ^ 0.1 ~ 2)或高(25 - 62 W m ~ 2)强度。在低照度的细胞生长在绿灯了藻红蛋白/叶绿素比率的两倍以上的细胞生长在蓝光。在三种红色macroalgae——纯粹elongata, Plocamium cartilagineum和Porphyra umbilicalis——来自海岸的岩石海岸马拉加(西班牙),在海水和孵化6个小时在不同光线下品质,Lopez-Figueroa和Niell(1990)发现,藻红蛋白的合成由绿灯优先刺激,和藻青蛋白的合成被红光优先提拔。在Cryptomonas物种隔离从深叶绿素层在西太平洋,Kamiya Miyachi
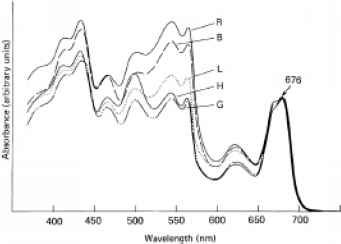
图12.8的影响光谱的光场在增长质量单细胞红藻的光合色素组成紫球藻属cruentum(许可,从雷和巴特勒(1980),植物生理学,65年,714 - 22)。光谱归一化到相同的吸光度值在676 nm。每个光谱信标签表示的光场细胞生长:R =红,B =蓝色,L =低强度白色,H =高强度白,G =绿色。
图12.8的影响光谱的光场在增长质量单细胞红藻的光合色素组成紫球藻属cruentum(许可,从雷和巴特勒(1980),植物生理学,65年,714 - 22)。光谱归一化到相同的吸光度值在676 nm。每个光谱信标签表示的光场细胞生长:R =红,B =蓝色,L =低强度白色,H =高强度白,G =绿色。
(1984)发现藻红蛋白/叶绿素比率更高的细胞生长在绿灯比细胞生长在蓝色或红色的光,相同的低辐照度(0.8 Wm ~ 2)被使用在每种情况下。我们可能认为这些色素的变化是互补的色适应:细胞增加,色素的比例最好吸收的光被暴露。
紫球藻属的细胞生长在高单色发光,然而,这个位置正好相反:细胞生长的藻红蛋白/叶绿素比绿灯还不到50%的细胞生长在蓝光。同样,雷和巴特勒(1980)发现,细胞的p . cruentum生长在高强度红(112 m ~ 2 s_1更易与光子)或蓝色(50 m ~ 2 s_1)更易与光子光对细胞生长在藻红蛋白/叶绿素比率的两倍高强度环保(99 m ~ 2 s_1)更易与光子光(图12.8)。在多细胞红色海藻Porphyra Yocum,眨眼(1958)发现,植物暴露于高强度蓝光(436 nm, 24 Wm ~ 2)十天包含更多的藻红蛋白和植物叶绿素比暴露在高强度的同时绿灯(546 nm, Wm 17.5 ~ 2)。
高强度单色光照明引起的色素成分的变化都伴随着植物的photo-synthetic特征的变化。Yocum和眨眼(1958)发现,海洋红藻,刚刚被收集或保存在绿灯十天显示在该地区光合效率低叶绿素的红色吸收带,光谱远低于下降的吸收光谱在650到700纳米之间。工厂一直保持十天在蓝光下,然而,在红色区域高效(以及蓝色区域显示出一些增加),行动和吸收光谱现在大约一致在650到700纳米之间。显然,从暴露在蓝光所导致(本身被叶绿素吸收)是提高光的利用效率被叶绿素吸收。
的本质photosyn-thetic系统中的自适应变化,发生在各种moderate-to-high强度、光谱选择性光政权已经被雷的详细研究,澄清了巴特勒(1980)对紫球藻属cruentum。仔细分析的吸收光谱和荧光行为他们已经能够到达结论有关的吸收特征,和之间的能量转移,这两个光系统在细胞生长在各种光谱成分的光。在红藻类叶绿素是主要的光色素光系统I和藻红蛋白在光系统II的主要色素。细胞生长在红色或蓝色的光会接收更多的激发能比phycoery-thrin叶绿素,,给定一个光合系统最初适应一个波长的广泛传播,将导致更大的能量输入光系统I比光系统II。为了确保均衡的光合系统的操作,必须增加的细胞吸收截面光系统II的相对光系统i他们实现的部分通过增加叶绿素藻红蛋白的比例,但也将更大比例的叶绿素光系统II。此外,雷和巴特勒发现这些红色或blue-light-grown细胞减少能量转移的概率从光系统II到光系统I,这有助于把激发能在光系统II。Porphyra增加红光的有效性,观察到Yocum和眨眼长期暴露海藻的蓝光后,我们现在可以合理的解释的比例增加叶绿素光系统II,因此一个更平衡的功能的光红灯。细胞生长在绿灯,如果他们最初适应波长的混合,得到更多的激发能在光系统II。他们的适应性反应是降低光系统II的吸收截面的相对光系统I通过减少藻红蛋白/叶绿素比值,并包括几乎所有的叶绿素光系统I: PS我叶绿素/ PSⅱ叶绿素在green-light-grown ~ 20 ~ 1.5相比redor blue-light-grown细胞。806此外,green-light-grown细胞表现出更大的能量转移的概率从光系统II我比细胞生长在红色或蓝色的光。
红色——或者blue-light-grown细胞藻红蛋白含量高于green-light-grown细胞在每个细胞的基础上,以及叶绿素比例。可能认为,尽管它可以把更多的叶绿素光系统II,使更多的藻红蛋白红光适应性反应,因为这不是一个有用的胆蛋白质几乎没有吸收超过600海里,所以不会收集红光子。然而,它可能是什么细胞的检测不是专门的光谱性质入射光,只是光系统I比光系统II接收更多的能量,所以它会用一个通用的色素补光系统II的增加。Red-dominated光场不自然发生在海洋环境中,我们不应指望红藻显示特定适应它们。
从雷和巴特勒看来,彩色的一般原则适应较高强度,可怕地限制,光场发生是光的组成和性质的调整等方式导致他们感到兴奋以相同的速度,从而高效的光合作用。布罗迪和爱默生的补色适应性观察(p . 1959) cruentum暴露于低强度的蓝色和绿色的光,因为它以相反的方式运作所以色素变化而言,必须有一个不同的基础。在低光强激发能的总供给的速度而不是光之间的失衡成为光合作用的主要限制。这可能是最好的策略,在昏暗的绿色或蓝绿色藻类生长光最好是让任何颜料将捕捉光线,将这些主要光系统II(胆蛋白质在这个光系统,反正)和转移的比例被吸收的能量光系统I。
色适应的蓝藻,涉及藻青蛋白藻红蛋白的比值的变化,是完全互补的类型并不奇怪。涉及的颜料都是胆蛋白质和两个饲料光系统II的激发能,所以问题不存在光之间的不平衡。增长的蓝绿色藻类Anacystis nidulans红色(1 > 650海里)光并导致叶绿素含量下降75%藻青蛋白含量变化不大。655年,我们现在可以解释这是细胞减少激发光系统I的点在与光系统II平衡。藻青菌的聚球藻属6301年,藻青蛋白/叶绿素比率增加红光(通过叶绿素吸收,主要是与光系统I)和减少黄灯(被藻青蛋白吸收,只与光系统II)。Manodori和梅丽莎(1986)解释这些变化代表实现平衡两光激发光系统的调整化学计量学:个人的实际天线大小光显然并没有改变。
继续阅读:问:0 10 15 20做0 3 10 15 Spfci造成phowuywvtfietro原始rrfl Cmachlj h mn C mc oflll 101 Cc h1
这篇文章有用吗?
读者的问题
-
莫妮卡6个月前
- 回复
-
牛顿1年前
- 回复
-
埃特1年前
- 回复
-
莱克斯1年前
- 回复
-
Mezan Tesmi1年前
- 回复
-
格蕾丝1年前
- 回复
-
杰拉德1年前
- 回复