蓝绿藻内的颜色适应
最明确的例子是在蓝绿藻(蓝藻)中发现的,它确实是在一个蓝绿藻物种中发生个体发生的色适应第一次被描述。Gaidukov(1902)观察到,在绿光下生长时,摇叶草呈红色,在橙色光下生长时呈蓝绿色:他将这些颜色变化归因于不同种类的色素的合成。Boresch(1921)指出,颜色的变化是由于类型的变化胆蛋白质由藻类合成:红色细胞主要含有藻红菊酯,蓝绿色细胞主要含有藻蓝蛋白。这种现象被Engelmann和Gaidukov(1902)认为是互补色适应的一个例子,由特定波段诱导的色素是吸收该波段的色素(藻氰菊酯和藻蓝蛋白的吸收峰分别在绿色(^565 nm)和红色(^620 nm)):“互补”是因为色素与诱导它的光具有互补色。
并不是所有的蓝藻都表现出颜色适应,在那些表现出颜色适应的蓝藻中,适应的形式也有所不同。Tandeau de Marsac(1977)将蓝藻细菌分为三组。在I组菌株中,胆蛋白组成不受其生长的光的光谱质量的影响,即它们缺乏颜色适应。在II组菌株中,只有藻红菊酯含量受光质影响,红光下含量极低,绿光下含量较高;在任何一种光照下生长的细胞中藻蓝蛋白含量都很高。在III组菌株中,这两种主要胆蛋白的合成受到光的光谱组成的影响:与前一组一样,它们的藻红菊酯含量在绿光下高,在红光下低,但它们的藻蓝蛋白含量在绿光下仍然很高,在红光下高1.6- 3.7倍。在检测的69株菌株中,I组25株,II组13株,m组31株
在I组和II组菌株中形成的藻蓝蛋白有两个多肽亚基,a和b。Bryant(1981)发现,在31个III组菌株中,有24个在绿光下形成的藻蓝蛋白也只含有两个亚基,但在红光下生长的细胞产生的藻蓝蛋白含有四个亚基,a1, a2, b1和b2。在绿光生长的细胞中,藻蓝蛋白的特殊亚基为a2和b1。这些元素总是存在的,科比称之为构成元素:另外两个,a1和b2,在红光下形成,他称之为诱导元素。因此,在这24个菌株中,红光诱导的藻蓝蛋白形成的增加是由不同种类的藻蓝蛋白亚基的合成组成的。不可能从数据中确定新的藻蓝蛋白亚基是否与形成不同的藻蓝蛋白物种(a1b2)n相关联,或者它们是否与两个组成亚基a2和b1聚合以形成杂交藻蓝蛋白。在其余7个III组菌株中,在红光生长的细胞中没有发现a1和b2藻蓝蛋白亚基:在这些情况下,额外的藻蓝蛋白可能只是由额外的a2和b1亚基组成。在toolypothrix tenuis中,Ohki et al.(1985)发现在绿光下的颜色适应导致藻红菊酯对藻蓝蛋白的一对一替代,因此藻胆体大小保持不变。
Ohki和Fujita(1992)发现,海洋蓝藻Phormidium sp. C86的细胞在红光下生长时呈深绿色,在绿光下生长时呈紫红色,这种颜色的变化是由于藻红菊酯的细胞含量大幅增加。红光下胆蛋白组成(摩尔基)为藻红菊酯13%,藻蓝蛋白64%,异藻蓝蛋白22%。在绿灯下,这一比例分别为82%、10%和8%。在绿光下生长的细胞中的藻胆体比在红光下生长的细胞大两倍。
Fujita和Hattori (1960,1962a, b)以及Diakoff和Scheibe(1973)在III组菌株之一Calothrix 7101(以前为toolypothrix tenuis)中研究了导致胆蛋白组成变化所需的光处理的具体性质。研究光对Calothrix 7101的控制影响是很方便的,首先在没有氮的情况下将细胞暴露在强光下24小时,耗尽细胞的胆蛋白,然后对所需的光谱质量进行短暂的光处理,然后将细胞置于有氮源的黑暗中,然后进行胆蛋白合成。在饱和剂量的红光照射后,细胞在随后的黑暗培养中合成藻蓝蛋白,但不合成藻红菊酯;在饱和剂量的绿光照射后,细胞继续合成藻蓝蛋白,但现在也合成藻红菊酯。如果交替给予红光和绿光处理,则随后的胆蛋白合成模式由最后一种光处理的颜色决定,即绿光或红光将各自逆转另一种光的效果
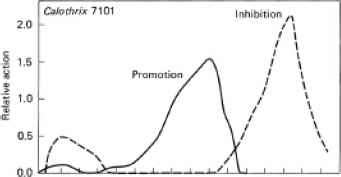
的光谱抑制藻氰菊酯合成的作用谱在红色约660 nm处有一个峰,在绿色约550 nm处有一个峰,在紫外约350 nm处有一个小峰(图12.7)
350 1d0米500 550 6d0小时700
WuvelcflUth \ NT我
图12.7行动光谱在蓝绿藻Calothrix 7101 (Tolypothrix tenuis)中促进和抑制藻红菊酯的合成(继Diakoff和Scheibe, 1973)。光处理后,在黑暗中合成的藻红菊酯(占总胆蛋白的比例)的量增加(促进)或减少(抑制)25%所需的辐射能(J cm-2)的倒数(归一化至500或680 nm后)与波长的关系。
350 1d0米500 550 6d0小时700
WuvelcflUth \ NT我
图12.7蓝绿藻Calothrix 7101 (Tolypothrix tenuis)促进和抑制藻红菊酯合成的作用谱(经过Diakoff和Scheibe, 1973)。光处理后,在黑暗中合成的藻红菊酯(占总胆蛋白的比例)的量增加(促进)或减少(抑制)25%所需的辐射能(J cm-2)的倒数(归一化至500或680 nm后)与波长的关系。
在紫外光下约350nm处有一个副峰。这些作用光谱的形状与光感受性色素系统本身本质上是胆蛋白相一致。Bogorad(1975)提出了“适应色素”一词,用于任何参与调节胆蛋白合成的光感受性色素。
Scheibe(1972)提出,这种控制色素类似于光敏色素,有两种相互转换的形式:一种(PG)有它的吸收最大在绿色和在绿光照射下进行光转换为另一种形式(PR),在红色区域吸收最大。在红光照射下,PR会被光转化为PG
绿色的高压
Scheibe指出,两种形式的色素都可能具有生物活性,PG促进藻蓝蛋白的形成,PR促进藻红蛋白的形成,或者只有一种形式的色素是活性的,它的存在或不存在决定了两条可能的分化途径中哪一条是被遵循的。Oelmuller et al.(1988)研究表明,在Fremyella diplosiphon中,绿光诱导藻红菊素信使RNA的形成,红光诱导藻蓝蛋白mRNA的形成。
虽然在光生长的细胞中,光的光谱质量会影响胆蛋白的合成模式,但光对于这些色素的合成并不是必需的:在黑暗的碳水化合物培养基上生长时,兼性异养蓝藻继续制造胆蛋白。在黑暗中培养III组这种菌株时,其胆蛋白组成与在红光下生长的细胞相似:高藻蓝蛋白,低藻红素Bryant(1981)发现,由Calothrix菌株7101和7601在黑暗中合成的藻蓝蛋白中含有可诱导亚基a1和b2,这种亚基在红光生长的细胞中发现,而在绿光生长的细胞中没有。无论所使用的接种剂来自于在红光或绿光下培养的培养基,情况都是如此。Bryant得出结论,不是暴露在红光下,而是没有绿光时,才会引发可诱导藻蓝蛋白亚基的合成。如果真的有一种光可逆的色素参与控制胆蛋白的合成,那么它可能是PG形式,它是由在黑暗中生长的细胞合成的。
继续阅读:真核藻类的色度适应
这篇文章有用吗?