系统发育色适应的证据
随着深度的变化,水生植物的色素种类在海水中变化显著,但在淡水中却很难识别,因此我们在这里将考虑海洋生态系统。我们首先要注意的是,在所有的海洋水域中,靠近水面的所有波段都有充足的光色适应它并没有告诉我们该地区不同藻类群的相对成功:例如,对波浪作用的抵抗,或者,对于上、次沿海形式,对偶尔暴露在大气中的抵抗,可能是更重要的因素。
虽然在所有水域中,总辐照度随深度呈指数递减,但光谱分布随深度变化的性质与水的吸收特性有关。在所有水域中,由于水本身的吸收,红色波段的辐照度会迅速减弱。在非常清澈无色的水域中,蓝色区域的衰减最小,随着深度的增加,水下光首先变成蓝绿色,然后主要是蓝色(图6.4a)。在含有大量黄色物质的水域中,蓝色区域和红色区域会迅速衰减,因此随着深度的增加,光越来越局限于绿色波段(图6.4b)。因此,在颜色适应的基础上,我们可能期望在不同类型的水中有不同的藻类分区模式,这实际上是事实。
三种主要真核藻类类群的深度分布可以用每一类群在一系列深度的生物量或物种数量来表示。这两种信息都很有趣,从我们目前的观点来看,第一种信息可能更有趣,因为它与通过初级生产不同群体的竞争成功更直接相关:相比之下,物种数量可以被视为绿藻、褐藻或红藻对在给定深度生长的问题找到的不同解决方案的数量的衡量,或者也可以被视为在该深度可以找到的绿藻、褐藻或红藻遗传信息的数量的衡量。这两种类型的定量分布数据似乎相当稀缺,不幸的是,在生物量的情况下尤其如此。
我们将首先考虑北半球温带的水域。对于瑞典西海岸(波罗的海的入口),Levring(1959)报道(尽管没有给出定量数据),绿藻的大多数成员出现在沿海和上副沿海地带,而Phaeophyta主要出现在海中滨海区下至亚滨海带中部(~15m),到处可见红藻真光区但在较低的次沿海带(15-30米)是主要的藻类群。北半球其他温带水域的分布与此大致相似,但似乎缺乏明确的分区。图12.1显示了在不列颠群岛的三个地点和北美东北部的一个地点,三个藻类类群的物种数量随深度的变化。绿色和棕色种类的数量随着深度的增加而减少。在一些地方,红色物种的数量先是随着深度的增加而增加,然后开始减少。在所有地点,绿色物种比其他两种物种少得多,棕色物种比红色物种少得多。在发现绿藻的三个地点(在锡利群岛的地点,绿藻显然没有或微不足道),它们深入到亚沿岸——大约到中部或稍深一点——但褐藻渗透得更深,红藻则更深。在所有情况下,红藻都主导着亚滨海的最低区域。Norton, Hiscock和Kitching(1977)测量了爱尔兰西南部一个海岬上较丰富的海藻生物量的深度分布,其数据绘制在图12.2中。绿藻在这个地点显然没有数量上的意义。从图中可以看出,棕藻几乎构成了整个亚滨海区的全部水下生物量。 Brown algal biomass consisted almost entirely of the large kelp Laminaria hyperborea: it reached its peak between about 6 and 10 m depth but decreased sharply below this, falling virtually to zero biomass at 18 m. Below 18m the vegetation consisted mainly of a comparatively sparse cover of red algal species. On a deep-water rock pinnacle in the Gulf of Maine (USA), Vadas and Steneck (1988) observed that kelps (mainly Laminaria sp.) were dominant from 24 m (pinnacle summit) down to 33 m depth. Foliose red algae were present through the kelp zone but extended further: they were dominant at 37 m and reached a maximum depth of 50 m. Crustose red algae became a significant component at about 37 m, and were the dominant algal type in the lowest region, fleshy crusts extending to 55 m and coralline crusts to 63 m depth. Green algae appeared to be absent from the rock pinnacle.
M Wttl苏格兰M Haw英格兰,美国*
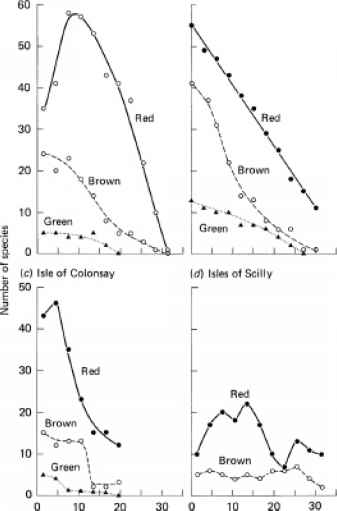
图12.1北半球温带水域底栖藻类区系分类组成随深度的变化。(a)苏格兰西部(Argyll and Ayrshire, 56-57°N).源自McAllister et al.(1967)的数据。(b)美国新英格兰(Maine和New Hampshire, 42-43°N),根据Mathieson(1979)数据绘制。(c)苏格兰内赫布里底岛(56°N)的Colonsay岛。来源于Norton et al.(1969)的数据。(d)英格兰锡利群岛(50°N),来源于Norton(1968)的数据。这些曲线显示了在每个深度发现的红色(-•-)、棕色(- o -)和绿色(…▲…)藻类物种的数量。曲线(a)、(c)和(d)由公布的每种藻类的深度分布得出。
深度i mj
图12.1北半球温带水域底栖藻类区系分类组成随深度的变化。(a)苏格兰西部(Argyll and Ayrshire, 56-57°N).源自McAllister et al.(1967)的数据。(b)美国新英格兰(Maine和New Hampshire, 42-43°N),根据Mathieson(1979)数据绘制。(c)苏格兰内赫布里底岛(56°N)的Colonsay岛。来源于Norton et al.(1969)的数据。(d)英格兰锡利群岛(50°N),来源于Norton(1968)的数据。这些曲线显示了在每个深度发现的红色(-•-)、棕色(- o -)和绿色(…▲…)藻类物种的数量。曲线(a)、(c)和(d)由公布的每种藻类的深度分布得出。
-
. Plotted from data of Norton, Hiscock and Kitch-ing (1977).")
- 图12.2北半球温带水域(Carrigathorna, lake Ine, Ireland, southwest, 51°N)中褐藻和红藻单位面积生物量随深度的变化。绘制自Norton, Hiscock和Kitch-ing(1977)的数据。
. Plotted from data of Norton, Hiscock and Kitch-ing (1977).")
北欧水域含有足够高水平的CDOM,以确保水下光线以绿色为主,519,636,美国东北部沿海水域可能也是如此。在这样的水域中,随着深度的增加,藻类的同化速率将取决于它在绿色(500-600纳米)波段的吸收量。绿藻在这一光谱区域的吸收相对较低,是最不利的,这可以解释它们对藻类群落的贡献(无论是物种数量还是生物量)都很小,而且它们的渗透深度最小。褐藻,由于岩藻黄质的存在,大量吸收在500至560 nm区域,通过大部分的沿海真光带保持自己。在真光带的下边缘,光谱分布变得相当狭窄,红藻,其胆蛋白质色素在绿色区域有它们的吸收峰,可以更好地争夺可用的光,因此占据主导地位。
现在我们将考虑在颜色非常低的沿海水域观察到的非常不同种类的藻类分布。泰勒(1959)在
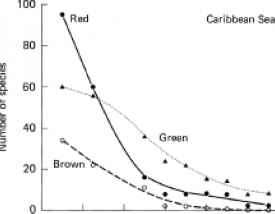
图12.3热带水域底栖藻类区系分类组成随深度的变化。根据Taylor(1959)的数据绘制。
图12.3热带水域底栖藻类区系分类组成随深度的变化。根据Taylor(1959)的数据绘制。
在加勒比地区,虽然红色、棕色和绿色物种在浅水中都有很好的代表性,但随着深度的增加,所有三组的物种数量都在逐渐减少(图12.3)。从比例上看,绿藻的减少最少,在亚滨海区域的低75%,绿色物种比红色或棕色物种更多。在太平洋也观察到类似的分布。在夏威夷沿海水域,绿藻种类虽然总体上数量较少,但渗透得与红藻种类一样深,而且比褐藻种类更深在埃尼威托克环礁泻湖,Gilmartin(1960)发现,尽管在65米以下的深度,绿色和红色藻类物种的数量相当(两者都大大超过棕色物种的数量),但在这一深度的所有站点上,绿色藻类似乎(根据视觉观察)在生物量方面占主导地位,在该深度,辐照度为表面的2%至4%。
地中海的藻类分布有一些相似之处。在科西嘉岛的一个岬角上,Molinier(1960)发现绿藻渗透到80米深处,在这以下被红藻所取代。Larkum、Drew和Crossett(1967年)进行了一项特别有价值的定量研究,研究了马耳他海岸垂直岩石表面藻类生物量的深度分布。他们测量了悬崖表面单位面积上三个分类组内藻类生物量的干质量,并将其作为深度的函数:结果如图12.4所示。下潜到大约10米深
-
. (Plotted from data of Crossett, Drew and Larkum, 1965.)")
- 图12.4地中海(马耳他,36°N)垂直岩壁上绿藻、褐藻和红藻崖面单位面积生物量随深度的变化(根据Crossett, Drew and Larkum, 1965年的数据绘制)
. (Plotted from data of Crossett, Drew and Larkum, 1965.)")
褐藻占主导地位,但它们对总生物量的贡献在这以下急剧下降。绿藻在15米左右变得显著,在20 - 60米(研究的最低深度)是群落的主要组成部分。红藻仅在30 m处成为重要组成部分,在45 m以下再次减少,比例和绝对数量都是如此。
Littler等人(1985,1986)使用潜水器对圣萨尔瓦多海山的藻类深度分布进行了详细的调查,在圣萨尔瓦多岛(巴哈马)以北6.5公里处,海山的顶部在海面以下约80米处形成了一个平方公里的平坦高原。高原地区和海山90米深的边缘主要是褐藻(%覆盖率),尽管存在各种各样的绿色、棕色和红藻。从90米到130米左右,有绿色和红色藻类,但以绿色为主,群落主要由4种钙质halalimeda属组成,特别是H. copiosa。从130米到189米,以壳藻红藻(Peyssonelia)为主,但在157米以上也有丰富的两种叶绿植物(至少在157米以上)。从189米到268米,优势物种是壳藻珊瑚红藻,但在210米以上出现少量的绿藻。
从这些不同的研究中可以清楚地看出,就藻类分布而言,无色的沿海水域和微黄色的沿海水域之间最显著的区别是,绿藻在前者中要成功得多。这正是在音色适应的基础上所期望的。正如我们所看到的,在这样的水域中,水下光场的蓝色波段特别丰富,绿藻就是在这个波段中进行大部分的光收集。除了在最深的地方,绿光和蓝光的比例都很高。一些种类的绿藻,由于含有类胡萝卜素虹吸-纳黄素(当与蛋白质结合时,在体内吸收500-550纳米区域),具有增强的绿光吸收能力。Yokohama et al.(1977)和Yokohama(1981)在对日本水域绿藻种类的调查中发现,在三个目(Ulvales, Clado-phorales和Siphonocladales)中,虹管黄素存在于深水物种中,而在浅水中生长的物种中则没有:这是颜色适应的一个合理的例子。在其他的一些目中,虹吸黄素存在于所有的物种中,甚至在那些属浅水区:在后者中,横滨提出虹膜黄素可能是来自深海祖先的进化遗迹。在海洋叶兰科(吊兰科)的14个物种中,O’kelly(1982)发现5种含有叶黄素(最丰富的高等植物/绿藻叶黄素)而不含虹吸黄素,4种含有虹吸黄素而不含叶黄素,5种同时含有叶黄素。只含叶黄素的物种主要分布在潮间带中上区,只含虹膜黄素的物种主要分布在潮下区,而同时含两种色素的物种主要分布在潮下区。
支持色适应理论的直接实验证据由Levring(1966,1968)获得。他测量了悬浮在瓶子里的绿色、棕色和红色藻类样本的光合速率,这些样本分别在瑞典和北卡罗来纳州海岸的混浊近岸水(绿色的透光率最高)和墨西哥湾流的清澈海水(蓝色的透光率最高)中进行了一系列深度测试。为了比较光合作用和辐照度随深度的变化方式,他使用了一个参数q,它可以看作是光合速率的垂直衰减系数与辐照度的垂直衰减系数的比值。如果某一特定类型的藻类随着深度的增加而越来越不适应光的光谱分布,那么q将大于1(即光合作用将比辐照度下降得更快);如果藻类能更好地适应深海底的光谱分布,那么q将小于1。在10米以下,对于绿藻来说,在混浊/彩色水中q为1.2 ~ 1.3,在无色水中q为^0.8,这表明它们更适合在颜色较浅的深水中进行光合作用。以红藻为例,在混浊/彩色水体中q为^0.8,表明适应能力随深度的增加而提高,在无色水体中q为~1.0,表明适应能力随深度的增加而变化不大。褐藻(Fucus)在有色/混浊水中q值为1.0,在无色水中q值为1.0以上或以下。
就浮游植物而言,也有系统发育的证据
色适应。在分层贫营养的蓝海海水中,叶绿素b集中在光以蓝色到蓝绿色为主的真光带底部附近,表明绿色绿植和原绿植的存在。在这些海洋区域通常发现蓝藻,如聚球菌,主要出现在海面附近,它们的高水平的光保护类胡萝卜素,玉米黄质,使它们能够耐受强烈的辐射。原绿藻,如原绿球藻,主要聚集在真光区底部的DCM中,在那里,二乙烯基叶绿素a的Soret带使它们能够有效地从主要的蓝光场中收集能量。237,1213,1412,72在近地表富含硝酸盐的水域中,硅藻占主导地位:它们的主要辅助色素岩藻黄质有效地收集大量存在于这些较浅深度的绿光。希克曼等人(2009)观察到,在季节性分层的凯尔特海广阔区域的水柱中,浮游植物类群存在显著的垂直梯度。色素组成和浮游植物吸收光谱表明不同的浮游植物群落在颜色上很好地适应了它们在水柱中出现的深度光场的光谱组成。Lazzara等人(1996)在热带北大西洋的一个中营养和贫营养区域,通过比较真光区底部辐照度的光谱分布(分别为500-530 nm和470-490 nm)与荧光激发曲线的峰值,发现浮游植物群落对主要光谱辐照度的互补色适应证据。在分层马尾藻海中,Bidigare et al.(1990)通过比较辅助色素的吸收峰与光谱辐照度最大值,发现了浮游植物对颜色适应的证据。
Pick(1991)研究了38个光学和营养状态不同的湖泊中不同色素类型的picocyanobacteria的分布。一些蓝藻picoplankton菌株只含有藻蓝蛋白和异源藻蓝蛋白胆蛋白色素,吸收峰分别在^620和650 nm处,而其他菌株也含有藻红蛋白,在^550 nm处吸收绿色。皮克发现,随着湖泊中光衰减的增加,这一趋势将伴随着水下光谱分布从绿色向红色的转变,因此含有藻红菊酯的皮青杆菌的百分比显著下降。Gervais et al.(1997)在德国Stechlin湖(Lake Stechlin)的分层水中,发现了一种含砷的低营养物质深层叶绿素最大值在10至15米的深度,盛行的光几乎完全局限在500至600纳米波段。DCM除了中心硅藻从上面沉降外,主要由含有藻红菊酯的皮青杆菌控制,它们在这些波长处有很强的吸收。
继续阅读:反对音色适应的证据
这篇文章有用吗?