光合作用时间的变化
这个问题的简短回答什么时候水生光合作用发生的是它发生时,在某种程度上,我们已经讨论了允许的各种限制约束。因此,理解光合作用时间的变化我们需要知道这些限制因素随时间的方式。
考虑日变化首先,晚上当然没有光合作用。光合作用开始在黎明和黄昏结束。整个水体的总光合作用大致遵循白天太阳辐照度的近似正弦变化。535659年、1337年仍然条件下可能会有减少光合速率单位体积表面由于附近光抑制。也,dinofla-gellates占主导地位的生物,他们向下迁移到低光强度可以降低光合速率单位体积表面附近的中间部分的一天。1360已经有许多报道的光合能力的变化(即光合作用在样本水体和暴露于饱和光)phyto-plankton人口底栖藻类白天(见,例如,Sournia的评论(1974),Raymont哈里斯(1980)和(1980))。这是稍后讨论(§12.6)。
在温带或北极/南极纬度,在水生光合作用有显著的季节性变化。这大约是,也是一个重要因素,植物生物量的季节变化。海洋浮游植物的详细叙述已经由Raymont (1980)。在冬天,浮游植物生物量和总光合作用都是非常低的,部分原因是太阳能irradiance1157但更低,因为由于降低温度和冬季的风暴,热分层现象失去和混合层的浮游植物是流传到超过临界深度净列光合作用(见§11.1)。在春天,增加表面的水加热产生的增加日常日晒,热分层现象,循环临界深度以下预防,这,结合事件的增加票面价值和营养物质在水中的可用性,导致大规模增加浮游植物生物量,通常的20倍,测量叶绿素。莱利(1942)发现的站在乔治海岸(缅因湾)在春季浮游植物的开花率增加(3月下旬至4月中旬)大约是成反比的深度混合区。他得出结论,之间的平衡垂直湍流和增加春天的辐射的影响决定了春天开花的开始(参见§11.1)。当垂直风混合匮乏或者较弱时,春天浮游植物爆发可以开始之前established.1373是热分层现象
一般春天开花后,下降到一个较低的水平的生物量和光合作用积分在夏天,也可能由于浮游动物放牧和养分的损失产生的混合层沉积的浮游动物粪便颗粒温跃层以下。在秋天,秋天在温度和风力强度的增加导致间歇性中断的热分层现象顺向运输营养物质从温跃层以下。因此,在海洋和大陆架水域在中间和高纬度地区,通常是有第二个浮游植物爆发和相关区域增加光合作用在秋季:这是终止当温跃层的破坏变得如此严重,分层消失,循环临界深度以下设置在冬天和生产力下降到较低的值。图11.7显示了全年光合速率的变化在北太平洋沿海水说明大弹簧,和小秋,峰值。734一般来说,随着纬度的增加,生长季节变得越来越短,春季和秋季浮游植物常常要合并成一个。
上面描述的温带的时空变化模式绝不是普遍的;它可以大大修改当地条件。卡特加特海峡(波罗的海),降低盐度(因此低密度)的上层保持全年分层,所以生产力是实质性的时刻,在冬天有所递减,由于减少了光的可用性。1297年沿海水域浅,混合深度很少或从不超过临界深度,全年生产也会居高不下。
-
.")
- 图11.7光合速率全年单位体积的变化在北太平洋沿海水(在Koblents-Mishke平均曲线推导出之后,1965年,从文献数据)。
.")
季节性变化在生产可能遵循温度循环如果光是non-limiting.1471的水很浅
Dandonneau et al .(2006)获得的数据的基础上,沿着12 trans-ocean邮轮海洋表面取样,在1999年到2002年期间,法国和新喀里多尼亚之间通过纽约、巴拿马和奥克兰证实可变性是最强的高纬度地区phyto-plankton生物量和人口结构有大量的地方季节性周期。春天盛开在北大西洋,叶绿素浓度通常> 0.5毫克m ~ 3,高浓度的fucox-anthin,硅藻的指标,而在夏天,叶绿素是低和人民都由原。球藻热带地区季节性变化和较低的特点是相对较高的二乙烯叶绿素a和玉米黄质(光保护类胡萝卜素)浓度。巴洛et al。(2007)使用色素分析浮游植物的季节性演替特征,和相关的叶绿素a和辅助色素的变化在亚热带水域的大西洋,印度洋和太平洋海洋在南半球。在低温度和辐照度条件下,光合类胡萝卜素是突出的,但随着温度和辐照度的增加有显著增加光保护类胡萝卜素的比例,这些最终成为色素池的最大组成部分,超过叶绿素,在最高温度和辐照度值。
随着卫星遥感海洋的颜色,它现在已经成为可以从太空监测季节性浮游植物的变化。
Stramska(2005)使用SeaWiFS年际变化数据检查的时间、强度和空间分布的浮游植物叶绿素在春夏装季节北极地区的大西洋,在此期间,1998年到2004年。每年发病的浮游植物爆发不同,演化和强度在一定程度上很大程度上由当地天气。开花的时间被推迟了几年有一个高速率的湍流动能(TKE)供应海洋今年3月,被风搅拌。今年4月,浮游植物的叶绿素和TKE供应率呈负相关。海洋中垂直混合与TKE供应的速度增加。佩雷斯et al。(2005)分析了浮游植物的季节性变化在赤道大西洋从1998年到2001年,使用SeaWiFS-derived叶绿素a浓度数据。他们使用了遥感在每月的间隔比较浮游植物叶绿素a值变化的位置(10°W)东部和西部(25°W)省。在两个地点叶绿素大约8月升至最高水平,但浓度达到两倍高在10°W 25°W·罗梅罗et al。(2006)使用SeaWiFS-derived月度叶绿素数据7年,1998年到2004年,研究季节性浮游植物年际变化的巴塔哥尼亚货架和货架,在南大西洋西部。45°以北,叶绿素在南国早春开花启动(Sept-Oct),而南45°的年代,开始在春末夏初开花(Nov-Jan)。生产率较高(> 4毫克的背影m ~ 3)比在开放海域(< 1.5毫克m ~ 3)。 Chlorophyll a concentrations in the northern mid-shelf sharply decayed in late spring, reaching their lowest values in summer (Feb-March), followed by a secondary maximum in early winter (June). All the highchlorophyll a regions were associated with well-defined fronts.
在旧金山湾的浑浊的河口水域,Cloern(1991)发现,每年春季浮游植物爆发有关密度分层导致从季节性增加输入的淡水河口。在较短的时间尺度,phytoplank-ton快速增长发生在小潮(低潮汐能,弱的垂直混合),但数量在下降大潮(高潮汐能量,强烈的垂直混合)。
在热带海洋水域没有季节性变化的水文状况,通常没有明显变化phyto-plankton生物量和光合生产。地区富含养分的深层水的季节性上涌,如非洲西海岸的部分,有一个伴随大规模增加浮游植物和光合生产。季节性河流流出的变化也会导致在热带沿海水域初级生产的季节性变化。
内陆水域在中间和高纬度地区的生物量和总光合作用显著的季节性变化。都是低在冬天由于辐照度下降(有时强调由冰或雪覆盖)和温度,在更深的水域,循环的浮游植物通过深度大于临界深度。在春天辐照度和温度增加,加上营养的可用性和热稳定的发作,导致浮游植物爆发和相关区域增加光合作用。的行为在其他季节往往是复杂和高度可变。生产可能会持续居高不下,或可能会有人口和生产造成的大范围波动浮游动物放牧、养分含量的波动由于大量的营养物从深水层与临时风致破坏的浅温跃层(海洋标准),或主要物种组成的变化造成水质的季节变化(如高pH值在夏末倾向于蓝绿藻而不是硅藻)。
离散的春天和秋天开花隔开一个夏天最低通常观察到湖泊,602、1120,但这时间模式更常见富营养的贫营养水体。870年图11.8展示了浮游植物生物量的变化,并通过半年每日浮游植物光合作用中滋育的康斯坦斯湖(Germany-Austria-Switzerland)。在一些热带湖泊如乔治湖,乌干达,可能没有季节性变化和初级生产和浮游植物生物量仍然很高。431另一方面,如乍得湖的情况下(乍得、非洲),有交替干燥和多雨的季节,温度的季节性变化,日晒和水位变化区域photosynthesis.795有关
底栖大型植物,被固定在适当的位置,并不直接与phyto-plankton不同,受季节性影响热分层现象的出现或消失。他们这样做,然而,显示季节性变化在光合生产的年度周期的辐照度和温度。图11.9显示了净光合速率的变化(单位面积的叶状体)的潮下带褐色海藻海带longicruris(海带)在新斯科舍,加拿大沿海水域通过的12个月里,一起价值总额的标准
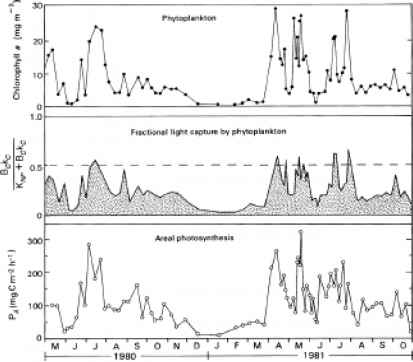
图11.8浮游植物生物量和光合生产的季节性变化。重绘从数据Tilzer(1983)中滋育的康斯坦斯湖(Germany-Austria-Switzerland)。中间的曲线代表的比例估计被phytoplank-ton事件相提并论。区域(积分)光合速率是中间的每小时平均4小时的一天。
图11.8浮游植物生物量和光合生产的季节性变化。重绘从数据Tilzer(1983)中滋育的康斯坦斯湖(Germany-Austria-Switzerland)。中间的曲线代表的比例估计被phytoplank-ton事件相提并论。区域(积分)光合速率是中间的每小时平均4小时的一天。
每天收到的海底(10米深度)藻类的生长,和水的温度。543年夏季光合速率最高,拒绝通过秋天大约11 - 12月刊的零,再次成为重要的冬季和春季末,然后在今年夏天早些时候,急剧上升。光合作用的时空变化对应大约辐照度的变化。通过多元回归分析,策划者et al .(1977)发现,61%的方差在日常光合作用可以通过辐照度占。温度的变化(从1.5到13°C在这些寒冷的水域)不占任何变化在日常光合作用的重要组成部分。温度是占56%的观察light-saturated光合速率的变化(以环境水温,
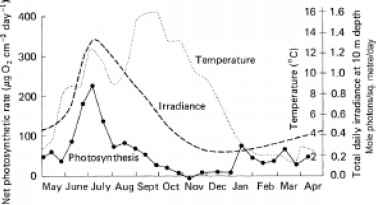
图11.9光合速率的季节变化的海藻植物longicruris和每日总辐照度(PAR)和水温度在10米深度在新斯科舍省,加拿大沿海水(圣玛格丽特湾)(从数据的孵卵的et al ., 1977)。光合作用原位测量是在流行的水的温度。每日总辐照度的图是通过绘制平滑曲线的数据点孵卵器等。
图11.9光合速率的季节变化的海藻植物longicruris和每日总辐照度(PAR)和水温度在10米深度在新斯科舍省,加拿大沿海水(圣玛格丽特湾)(从数据的孵卵的et al ., 1977)。光合作用原位测量是在流行的水的温度。每日总辐照度的图是通过绘制平滑曲线的数据点孵卵器等。
回到实验室),但因为大多数时候植物原位将光合作用subsaturating辐照度值,温度对饱和率的影响是无关紧要的。
教徒和里德(1979)研究了海草,伊南极光,生长在浅水——从最低点的深度3米以下这个黑客港,澳大利亚(34°S)。图11.10显示了相对增长率的变化(mg C g - 1普通人级,这应该与光合速率密切相关),和水温度在12个月期间。增长率被发现与水温密切相关(r = 0.79)。在墨西哥海湾北部浅海草床,另外,优势种的季节生长周期,Thalassia testudinum,关联更紧密地与水温比太阳能辐照度。840年很可能在这些浅,有很好的照明环境,光强度都达到或接近饱和的时间,温度成为限制因素。
Hanisak(1979)进行了综合和实验室研究领域的增长模式siphonous绿藻松藻属脆弱,生长在浅水(平均深度1.5 - -2.3米以下的意思是最低点)(41°N)东北部海岸的美国。增长的主要时期(干物质积累,因此光合作用密切相关)
-

- J_I_I_I_I_I_I_I_I_I_L


小君吗?7月Auq 9月!Oci Hw 12月1573年1月1977年2月4月梅
月
图11.10季节性变化的相对增长率海草伊南极光,地表水温度,黑客港、新南威尔士、澳大利亚(由教徒和许可,里德(1979),水生植物,7,173 - 83)。
小君吗?7月Auq 9月!Oci Hw 12月1573年1月1977年2月4月梅
月
图11.10季节性变化的相对增长率海草伊南极光,地表水温度,黑客港、新南威尔士、澳大利亚(由教徒和许可,里德(1979),水生植物,7,173 - 83)。
从春末(环比)到初秋,7月份的最大速率。每天似乎总辐照度水平高于饱和增长从4月到8月/ 9月温度是主要的限制因素,但是在秋天,气温仍然很高时,减少光水平限制了经济增长。冬天似乎温度和光线都是限制。
季节性变化在一个为期两年的研究在普吉特海湾底栖生物macroalgae /海草社区(华盛顿、美国),汤姆·奥尔布赖特(1990)发现,在水温最好似乎关联站股票,太阳辐照度变化似乎触发生物量积累在春天发作,和在秋天枯萎的。期间光和温度限制时,水中的硝酸盐水平增长成为限制因素。
在一些沿海地区的减少水下光可用性在冬天由于降低太阳能高程和短天显著增加,进一步加剧在垂直的光衰减造成水浊度增加激起造成的沉积物在冬季风暴。Banyuls-sur-Mer(地中海西部),从最小值约为0.075 Kd上升1夏天最高大约0.19 1在冬天,1447和Helgoland(北海)另外,从十月至三月水透明度低得多比其他year.833
这些数据表明初级生产的初步概括在深水海洋大型植物有限光可用性,但在浅水通常限于温度,除了在高纬度地区的光成为限制在冬天,和时态的变化主要生产的是在很大程度上的季节性变化的函数这两个物理参数。很可能大型植物光合作用(除了在深水中),此外,时时处处受到二氧化碳的可用性,但这并不随着季节而变化。增长,因此光合作用,还取决于可用性养分的水,尤其是硝酸盐,在高纬度地区不同在季节性的基础上高在冬天但大大减少在春季浮游植物爆发。这是更少的限制因素为许多大型植物比浮游植物,然而。海草,例如,源自植物,可以从沉积物中获取营养生长。在夏天,当水中的硝酸盐浓度较低,海藻(褐藻类)继续光合作用和碳水化合物积累储备以备后用,在冬天。543在热带地区我们可以先验预期相对较少的大型植物光合作用的季节性变化除了哪里有水文状况的变化,例如季节性河流流出的变化有关。
季节性变化的程度在海洋大型植物光合作用受到内部的影响,以及外部的变化还不清楚。国王和施拉姆(1976)在他们的研究的许多绿色,棕色和红色藻类物种在波罗的海,发现,例如,在冬天,光补偿点低比在一年中的其他时候,light-saturated光合速率(每克干质量)在任何物种明显不同(但不是以同样的方式在所有物种)根据季节。然而,他们的测量在所有情况下进行盛行的季节水温(年)期间,当然,不同的,所以他们无法得出季节性是否适应只是直接响应温度变化或其他因素是否参与。季节性变化的一个例子在光合作用由内部生理变化引起的,但是,提供的海草伊大洋洲。画(1978,1979)发现,在植物的物种生长了马耳他的光合速率(单位面积的叶)在夏天明显低于春季价值下降,而在另一种海草物种,Cymodocea nodosa,附近的增长,光合速率仍然很高。p .大洋洲光合作用的下降似乎是由于衰老(夏天叶叶绿素含量显著下降),可能引发的昼长变化。
在新鲜水,底栖大型植物初级生产的典型模式在温带地区生物质积累不发生在冬天,它开始在春天的太阳辐照度和温度的增加,并上升到其最大速率(g C或每平方米每天底)干物质在初夏。在macrophyte-dominated Gryde河(丹麦),凯利et al。(1983)发现,每日在一年整个植物群落的初级生产力密切关注每日日晒。因此,据推测,光的饱和,有一个明确的趋势在某一天生产的效率与白天增加辐照度降低。光和温度的相对重要性决定的季节性模式淡水大型植物光合作用还不是很清楚。我们可以合理怀疑,它主要是一种深度,温度在浅,更重要的变量,光在更深的水。在夏末和秋季,积累的速度下降的原因包括总体恶化的活力植物由于疾病、损伤和过度放牧的温度。1456的pH值在夏天晚些时候,由于高浮游植物光合作用,也可以对一些大型植物有害。大型植物可以推出以来从沉积物中营养物质增加,他们的生产力不可能有限的季节性变化水平的磷和无机氮在水里。图11.11显示了初级生产的季节性变化速率在混合在一个大型社区加拿大的湖.830内部以及外部的变化可能影响光合作用的季节性变化:淡水大型植物的光合能力通常是低在冬天和春天高。1455光合能力的变化可以影响植物光合作用的速率在浅水的饱和强度,但是对植物的后果很小在深水subsaturating强度。
继续阅读:系统发育色适应的证据
这篇文章有用吗?
读者的问题
-
斯蒂芬。2个月前
- 回复
-
Emppu4个月前
- 回复
-
alistair肯尼迪6个月前
- 回复
-
不7个月前
- 回复
-
alfrida8个月前
- 回复
-
玛丽8个月前
- 回复
-
托马斯。8个月前
- 回复