行为反应,紫外线辐射
行为反应通常辐射随波长。一些
-
 green (490-560 nm) and (b) ultraviolet (350-380 nm) wavelengths. Note the bright background in the UV image that silhouettes fish strongly, even against the reef. [Taken from Losey et al. 27.]")
- 图3。同时在拍摄的图像(a)绿(490 - 560 nm)和(b)紫外线波长(350 - 380海里)。注意明亮的背景在紫外图像轮廓鱼强烈,甚至对礁。(取自一丁点它们et al . 27。)
 green (490-560 nm) and (b) ultraviolet (350-380 nm) wavelengths. Note the bright background in the UV image that silhouettes fish strongly, even against the reef. [Taken from Losey et al. 27.]")
图2。的比例对紫外线a(380海里)光合有效辐射(PAR, 400 - 700海里)的W m ~ 2。期间收集的数据是2001年的夏天,紫外线辐射仪(模型Biospherical老爸- 521)位于Lacawac保护区在波科诺Mts, PA,美国(41.23 N, 75.21 W)。日出(5.27 h),日落(20.37小时)2001年6月17日是用垂直的虚线。日出日落2001年7月8日发生在5.36 h和20.37 h,分别。朦胧的时期,紫外线:PAR比率较高,因为光场主要是由天窗(见14.1节)。随着太阳的高度增加,票面金额的增加,由太阳辐射的光场。注意,增加类似UV-A-to-PAR发生在云经过太阳的补丁。这是17.0 - -19.0 h之间显示2001年7月8日。
17-jun——8-Jul诱发积极的趋光性或运动对光源波长而其他波长诱导负向光性或者运动远离光源。对于许多物种,暴露在紫外线辐射(280 - 400 nm)短波长(即可见。,蓝色光,400 - 440 nm)光引发负趋光性。这些消极phototactic或回避行为对应的波长也被认为是潜在的破坏性或致命(11、16)。在能动的有机体,水平和垂直方向的运动一直在避免暴露于有害辐射(28-31),而在不能动的有机体行为表现出[32]。许多生物,如海葵、海胆、海参,以贝壳、岩石、和其他材料在高峰时期的辐照度。隐藏在洞穴和岩石和大型植物床,见许多两栖动物和幼虫鱼类,也有助于生物减少暴露于有害辐射。
14.3.1实验室实验
迄今为止,行为反应破坏光主要是研究在实验室利用人工紫外线辐射来源。实验已经进行了各种各样的生物从淡水和海洋系统占据所有的营养水平。营养水平较低,浮游植物和原生动物都表现出负趋光性紫外线辐射。例如,单个细胞内垫的丝状海洋蓝藻Micro-coleus chthonoplastes被证明迁移到更大的深度,以应对uv - b增加曝光[28]。红色淡水纤毛虫赭纤虫日本血吸虫了逆向游泳当暴露在紫外线的波长范围,开始向前游泳当暴露于可见光在580 nm [33]。调查是很重要的在这些低营养级生物的行为因为他们对紫外线辐射的反应可能直接或间接地影响反应的高营养水平。
实验室实验清楚地表明的波长入射辐射是一个重要的行为暗示浮游动物。某些淡水枝角目动物变得更加激动和消极phototactic蓝色光的存在,但保持冷静,积极phototactic红光[29]。这些“颜色舞蹈”枝角目并假设暗示浮游动物高浓度藻类食物,一般过滤掉短的波长大于长波长(即。red-dance“保持个人,“blue-dance”促进徘徊)。然而,它也表明,舞蹈的模式可以解释的昼夜垂直迁移模式。最近与单色辐射的研究已经证明水蚤麦格纳是积极phototactic可见光(421 - 600 nm)和负面phototactic紫外线辐射(260 - 380 nm)和最大灵敏度为340 nm[34],桡足类也表明在实验室紫外线回避行为。在小实验附件检查水平运动,淡水cyclo-poid独眼巨人serrulatus发现避免暴露于紫外线辐射(280 - 320 nm) [10]。这项研究还指出,紫外线与紫外线耐受(即行为反应相关。UV-sensitive生物避免紫外线的照射,见14.3.4节)。避免紫外线辐射行为也在海洋中发现海胆状的幼虫Dendraster excentricus暴露在人工紫外辐射源(315 - 700 nm) [30]。
某些stream-dwelling生物已被证明是负面phototactic紫外线辐射在实验室实验缩影。大型无脊椎动物,居住或饲料在顶部或两侧的岩石,如蜉蝣的幼虫阶段,毛翅蝇,和黑蝇,表现出漂移增加更多的阴影区域暴露在紫外线辐射增加[31]。漂流60 - 70%的紫外线屏蔽控制。
有趣的是,深海甲壳类动物对紫外线辐射的反应行为。拴在个人的oplophorid虾Systellaspis debilis应对变化的环境紫外线辐射的投手,改变游泳速度和移动他们喂养附件[35]。可能的解释行为反应紫外线14.4.2节讨论在深海甲壳类动物。
对于一些生物,完全暴露在紫外线辐射抑制运动。例如,暴露于人工紫外线后,面盘幼体和斑马贻贝的post-veligers Dreissena polymorpha停止所有游泳和爬行运动。然而,暴露于紫外线a和可见光没有影响行为[36]。类似的延迟趋光性是绿藻指出团藻球菌暴露在人工和太阳紫外线辐射[37]。
尽管这些研究提供有价值的信息关于有机体的反应在不同波长的辐射(即,行动光谱),他们不告诉我们动物如何应对自然的太阳辐射水平。人工灯通常不完全复制太阳光谱。紫外线灯通常有更大的输出在uv - b与太阳光谱范围。为了补充对紫外线a和可见光、紫外线a和酷白灯被用于实验室设置,这些灯经常少产出比太阳辐射对紫外线a和可见的范围。这些灯的总强度的能量或量子可能类似于太阳辐射,但(即光谱组成有很大的差异。,偏向较短的波长)。太阳模拟器是最接近太阳的强度和光谱输出复制;然而,这些工具很贵,只有照射面积小,只有少数实验室使用。
14.3.2领域使ts
很少有现场研究检查行为反应的生物自然太阳辐射。这些研究的困难之一是确定行为是否以应对紫外线辐射或可见光。高紫外线系统也高可见光系统,我们知道这两个潜在的破坏性[1 8 38]。此外,许多动物都有一个单独的套房的反应不同程度的可见光。通常情况下,实验附件在紫外线辐射透射率不同的材料建筑而成。常用材料传输完整的太阳辐射包括聚乙烯、石英、和丙烯酸塑料如OP-4 (CYRO产业)和UVT (Spartech公司以前Tow-nsend / Glasflex),所有的这些都可以是昂贵的。防紫外线材料包括聚酯薄膜®D和丙烯酸塑料如OP-2 (CYRO产业)和树脂玻璃®。虽然这些材料不同紫外线透明,他们有类似的幻灯片在可见范围内。因此,使用这些材料的组合,行为和/或生存可以检查在完整的太阳辐射的存在,没有紫外线辐射,在uv - b和a的缺失
辐射。重要的是要注意,这些实验不提供有关信息对单一波长的反应;相反,他们检查删除特定波段的影响(即。、uv - b或uv - b和a)。因为他们删除整个波段紫外线屏蔽材料也不可避免地改变总辐照度,从而影响最终的结果。然而,紫外线之间总辐照度的差异——透明、防紫外线材料通常是不到10%。最后,除了石英,它非常昂贵,而且很难制造,通常将材料往往阻止紫外线的重要部分(25 - 50%)。
,太阳能uv - b已被证实能抑制活性和定向运动在真核鞭毛虫等浮游植物,蓝绿藻或蓝藻,滑翔绿藻[39]。当活性被破坏时,浮游植物在被暴露的风险更大的光强度,这可能会导致色素漂白;或者他们可能接触到光强度降低,这可能会导致光合率的减少。暴露在增加或减少辐照度还取决于混合过程以及浮力的单个细胞(见第4章)。
最近的领域的研究也报道,浮游动物展览避免紫外线辐射。垂直回避反应的第一个证据水蚤太阳紫外线辐射是最近出版的[40]。在完整的太阳辐射,d . pulicaria迅速后裔的表层水(1.5米)高紫外线的湖。没有紫外线和较短的波长的紫外线a辐射(< 380海里),大多数d pulicaria留在表面水域。因此,一个更强大的负面phototactic响应中检测出的存在比没有紫外线辐射的紫外线辐射。负phototactic行为也被观察到具有d·卡托巴人居住人口的高紫外线湖位于波科诺Mts, PA,美国(图4)[41]。实验进行的这项研究表明,在某些情况下,d·卡托巴语实际上游泳对地表水缺乏紫外线辐射,尽管可能存在的鱼它激素对水蚤(图4)。这些字段的结果是由规模较小的实验在实验室(42、43)。
虽然水蚤经常表现出偏爱的地表水缺乏紫外线辐射的情况下,响应变量,平均深度水蚤增加没有紫外线辐射[41]。这个变化的原因是未知的。一种解释是,辐照度实验之间的不同。尽管这项研究没有设计具体实验测试浮游动物对辐照度的反应,初步观察表明,随着辐照度的增加,水蚤回应增加负趋光性。因为丙烯酸用于构造防紫外线列传输一些长波长紫外线a(50%透光率在384海里),这可能是一个回避应对长波长紫外线a光或可见光[40]。其他物种,如淡水桡足动物Diaptomus nevadensis,海洋桡足动物Acartia tonsa,水蚤类的水蚤麦格纳,和hydromedusan Polyorchis penicillatus也变得消极phototactic辐照度增加,紫外和可见的范围(1、38)。
的harpacticoid桡足动物Tigriopus californicus,住在浅潮
向下
向下
-

- 向上

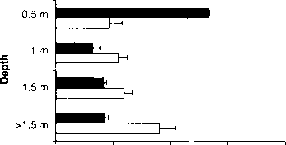
0 20 40 60 80
%的人
图4。比较的向下和向上迁移水蚤卡托巴语的存在和没有紫外线辐射。有三个将列和三个UV-opaque列。每一列是悬浮在地表以下10厘米的吉尔斯湖,位于宾夕法尼亚州东北部高紫外线的湖,美国。向下的实验进行了2000年7月14日和2000年8月2日上升。平太阳辐照度测量与李LICOR模型- 200 sa中午太阳附近的日射强度计实验时(1300 h)。平太阳辐照度等于659 Wm于2000年7月14日~ 2和694 Wm ~ 2 2000年8月2日。
0 20 40 60 80
%的人
图4。比较的向下和向上迁移水蚤卡托巴语的存在和没有紫外线辐射。有三个将列和三个UV-opaque列。每一列是悬浮在地表以下10厘米的吉尔斯湖,位于宾夕法尼亚州东北部高紫外线的湖,美国。向下的实验进行了2000年7月14日和2000年8月2日上升。平太阳辐照度测量与李LICOR模型- 200 sa中午太阳附近的日射强度计实验时(1300 h)。平太阳辐照度等于659 Wm于2000年7月14日~ 2和694 Wm ~ 2 2000年8月2日。
池,发现总在中午池的阴影区域,但没有偏好在黎明和黄昏[44],这些相同的作者用实验证明t californicus响应更多的紫外线比可见辐射和表明他们可能拥有UV光感受器。
小溪无脊椎动物也被指出消极应对紫外线辐射。黑蝇幼虫似乎表现出昼夜移民或移民UV-exposed流渠道高峰时期,辐照度但回到UV-exposed地区随着辐照度水平降低[45]。在流实验免受紫外线照射,然而,幼虫仍在流渠道。幼虫被允许自由移动和治疗之间的平均幼虫密度UV-shielded频道的161 - 168%大于那些UV-exposed频道。
黄色产卵深度的差异,如刺蛾,在高位低紫外线湖也表明黄鲈避免紫外线照射。产卵深度报道中更深层次的高紫外线湖(值= 3.2 m)相比,低紫外线湖(值= 0.4 m) [46]。此外,黄鲈蛋在孵化中的每个湖的表面改性相互移植实验。鸡蛋被暴露于太阳辐射,从紫外线屏蔽,或者保持在黑暗中。高紫外线的湖,所有鸡蛋死于光治疗,但存活时间长(2 - 4天)紫外线保护鸡蛋的治疗。此外,那些来自高紫外线收集的存活时间比低紫外线湖湖。大多数(> 96%)孵化鸡蛋低紫外线光疗法的湖以及两个湖泊中幸存下来的黑暗控制孵化。类似的结果,使用一个类似的实验设计,据报道蓝鳃太阳鱼Lepomis machrochirus中位数中被观察到的更深层次的嵌套深度高紫外线湖相比,低紫外线湖[47]。
它更难以在公海执行行为实验。方法是观察生物的分布与光环境结合实验室实验研究紫外线宽容和趋光性。海鞘和海胆都表现出避免紫外线。孤独的海鞘类的分布Corella inflata随接触直接太阳辐射,尤其是紫外线照射,人口明显缺席没有阴影的区域[48]。实验证实,紫外线辐射是致命的生活史阶段的c . inflata年轻的阶段是最脆弱的。此外,没有一个生命阶段具有防紫外线光保护化合物。的海胆Arbacia punctulata和排泄殖腔被证明是负面phototactic明亮的太阳辐射,但积极phototactic白光[49]。这些数据符合echinoplutei迁移到更深的深度的观察辐照度的水柱在高峰时期(30、50),但是这种反应也可以避免捕食者等其他因素有关。海胆Strongylocentrotus droebachiensis阴影或覆盖本身紫外线照射,特别是在响应与紫外线或uv - b和a [32]。覆盖的行为也证明增加紫外线照射的强度增加。 In some sea urchin species, covering behavior has been observed to vary在白天在高峰辐照度,最大的反应[49,51]。
14.3.3与紫外线宽容,色素沉着,andphotorepair
行为对紫外线辐射的反应似乎与紫外线耐受(即有关。,定义为一个有机体的总和的光(色素)和photorepair功能)。例如,经过一段时期的高紫外线,有机体占据湖贾尔斯的地表水,紫外线高湖在波科诺Mts, PA,美国被发现更UV-tolerant白天比居住在更深的水域[12]。实验室实验和介形亚纲动物腺介虫sp.证明这个物种对uv - b照射高度宽容实际上显示行为偏好或正向光性对紫外线辐照度[10]。在相同的研究中,原生动物草履虫蛹也被证明是高度宽容和积极phototactic与紫外线辐照度[10]。
的光谱趋光性的桡足类已被证实能依靠色素沉着。在可见光谱中,Diaptomis nevadensis游泳速度快在蓝光比红光[38]。此外,色素减少人对波长的变化比色素个人[52]。类似的结果也出现在melanized水蚤在紫外光谱[43]。
继续阅读:紫外线视觉和视觉感受
这篇文章有用吗?