模式的DNA损伤积累和水生生物修复
9.5.1从实验室研究证据
在水生生物的DNA损伤诱导的很多信息都是基于人工紫外线辐射下孵化实验。不幸的是,这样的生态相关性研究限制,主要是由于人工之间的光谱差异(灯)和太阳辐射来源。结果,uv - b的影响可能高估了即使加权紫外线辐照度是现实的。另一方面,实验可能提供有价值的信息对物种的具体差异紫外线脆弱(见9.6节),潜在的修复,或各种环境因素对紫外线脆弱性的影响。Joux等。[68]清楚地显示,海洋细菌物种特定DNA损伤积累,主要是由于不同的光致复活作用的潜力。早期研究显示潜在的CPD积累和蓝藻和硅藻photo-enzymatic修复能力[9,55岁,69 - 71]。pak等。[72]6 - 4页和CPD积累在海洋红macro-alga Palmaria palmata并显示成功克服uv - b的光致复活作用压力的重要性。另一项研究关注海洋红藻[73]相关的uv - b模式的脆弱性CPD积累和修复。他们发现滨海红藻更不容易遭受CPD积累与sub-littoral物种和相关主要产能photorepair uv - b的脆弱性。
在海洋生物没有直接测量DNA损伤诱导,许多研究集中在海洋生物光致复活作用的潜力。汉和实物地租[74]证明了光致复活作用的孢子体由蓝光在年轻的几个褐色海藻。他们强调,可见光导致复苏紫外线的伤害在这些生物,否则造成高死亡率。Huovinen等。[75]研究了紫外线辐射的影响在早期发育阶段的巨型海藻Macrocystis pyrifira暴露孢子萌发前后紫外线。他们发现,核分裂和易位在游动孢子萌发为紫外线辐射最敏感。此外,复苏发生在光,可能由于光致复活作用受损DNA在游动。Huovinen et al。[75]的结论是,短曝光与紫外线可能会扰乱或延迟的发展和招聘巨头海带配子体通过抑制他们的核分裂和易位。Coohill和-迪尔岭[76]发现了一个清晰photoreactivating波长之间的关系和生存的延长水霉菌Blastocladiella emersonii。此外,他们还发现,温度主要影响光致复活作用,高温导致更高的生存的孢子。
CPD形成和潜在的光致复活作用已经描述了鱼细胞系,活鱼和鱼类胚胎[77 - 80],Kouwenberg等。[81]认为紫外线引起的死亡率在海洋浮游动物和鱼可以归因于DNA损伤。生物加权函数蛋死亡率在大西洋鳕鱼高相似之处的波长依赖DNA的DNA损伤诱导的解决方案,如发表的Setlow [33]。他们也证明了降低鸡蛋的死亡率photoreactivating波长的存在。海洋浮游桡足类的Acartia omorii,鸡蛋比老年生活更敏感阶段[82]。UV - b诱导损伤与增强PAR同时缓解下辐照度,表明photoenzymatic修复中起着重要作用在确定紫外线压力在这些生物。Kouwenberg等。[81]的结论是,年轻的阶段可能主要受事件影响紫外线辐射。Zagarese et al。(83)展示了物种特定photoenzymatic修复效率为各种浮游动物物种。海斯等。[84]测量两栖动物蛋死亡率作为光裂合酶活动的函数。他们发现光裂合酶改变100倍的鸡蛋10两栖动物物种。此外,孵化成功与光裂合酶活性[84]密切相关,
很明显这些实验室研究光致复活作用可能是一个关键过程在确定最后的uv - b在水生生物反应。大多数生物调查photorepair潜力,即他们展示photoenzymatic活动“光致复活作用光”的存在。然而,这种潜在的光致复活作用可能不是完全利用水生生物在他们的自然环境。首先,光谱条件水是高度动态的,可能并不总是支持photoenzymatic过程。例如,在更大的深度,对紫外线a维修或票面可能仍低于阈值。同时,化学和能量要求修理可能并不总是足够的。因此,原位积累和修复模式需要进一步阐明photorepair的有效性及其影响水生生物的DNA损伤积累。
9.5.2深度相关的CPD模式,一昼夜的周期和混合效果
在深海生物,uv - b照射在很大程度上取决于混合现象。由于风引起的垂直混合,生物在上层水体上下移动。因此,生物体的不同波长的强烈波动,将很难确定(见第4章)。关于DNA损伤,诱导修复率也将在混合,因为两个过程强烈波动,但以不同的方式,波长的依赖。垂直混合的后果对DNA损伤积累和修复可以使用模型评估等提出的Huot et al。[85]。他们总辐照度、混合和生物模型预测净混合层的DNA损伤水平。模型的预测通常是符合测量DNA损伤水平。模型表明,混合层深度的强烈影响的数量和分布混合层的DNA损伤,但DNA损伤的平均数量在整个透光层只有小受混合的影响。此外,模型的计算表明,光致复活作用中起着重要作用在防止海洋bacterioplankton快速CPD积聚。
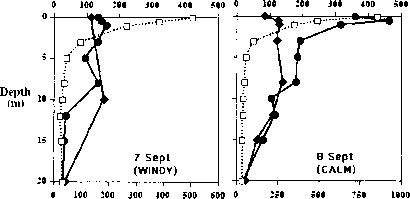
最近,许多原位观察紫外线有关海洋生物的DNA损伤积累出版。最小的生物居住在水生生态系统是病毒。病毒裂解可以鞭打放牧导致死亡率一样重要的海洋细菌社区。UV-induced病毒传染性可能的破坏,因此,降低病毒介导死亡率bacterio——浮游植物。病毒已被证明在海洋水域DNA损伤积累,从而减少病毒的传染性。cpd和6 - 4页显示一个明确的深度模式在墨西哥湾的[12](6 - 4页配置如图5所示)。在被观察到了类似的破坏模式bacterio-plankton在南大洋(Jeffrey et al .,未发表的结果)。在他们的研究Weinbauer等。[12]描述了感应cpd和6 - 4 PPs在病毒隔离,孵化出了几个水深度,而且在自然病毒社区抽样在一天结束的时候(集中于特定的深度)。以PPs和cpd显示明显相似的分布与深度。6 - 4页的比例相对于总DNA就会受到明显损害平均3.1%的自然病毒社区在墨西哥湾。一般高损伤水平固定隔离与自然相比原位样品表明混合率最小化photodamage积累在病毒DNA。 This conclusion was further supported by the CPD depth profiles of bacterioplankton from the Southern Ocean [86] and the Northern Gulf of Mexico [11,87] (Figure 6). During calm seas, damage was highest in the surface, decreased with depth and could be detected down to 10 m in the Gulf of Mexico. On moderately mixed days, however, no net accumulation of damage was observed, not even at the surface. In the Gerlache Strait, Antractica, damage was observed to accumulate below 20 m on a calm day. When seas approached 1 m, however, DNA damage at the end of the day was less than it was at sunrise [86]. It was postulated that damaged cells may be moved to deeper waters by mixing, where rates of photoreactivation may outweigh rates of UV-B induced damage, resulting in net decreases of DNA photoproducts.
虽然一定程度上(参见9.6节),微藻也积累
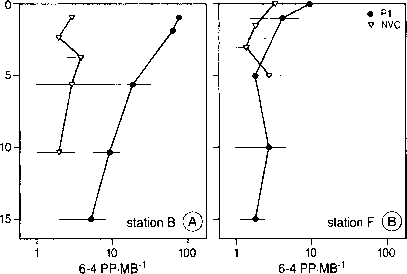
图5。深度资料6 - 4页的噬菌体的DNA,在两个电台(站B和F)在墨西哥湾。满圆:PWH3a-Pl噬菌体分离,孵化整个每日段在不同水深处。开放的三角形:自然病毒社区集中从特定的深度在一天结束的太阳能。(重绘Weinbauer et al。12。)
图5。深度资料6 - 4页的噬菌体的DNA,在两个电台(站B和F)在墨西哥湾。满圆:PWH3a-Pl噬菌体分离,孵化整个每日段在不同水深处。开放的三角形:自然病毒社区集中从特定的深度在一天结束的太阳能。(重绘Weinbauer et al。12。)
环丁烷二聚体/ Mb DNA (bacterioplaiikton)
环丁烷二聚体/ Mb DNA (bacterioplaiikton)
环丁烷二聚体/ Mb DNA (Dosiineters)
图6。CPD深度剖面的细菌组合和测试仪墨西哥湾北部,以6.30 h和19.00 h。左面板:1994年9月7日。右面板:1994年9月8日。(重绘Jeffrey et al。11。)
环丁烷二聚体/ Mb DNA (Dosiineters)
图6。CPD深度剖面的细菌组合和测试仪墨西哥湾北部,以6.30 h和19.00 h。左面板:1994年9月7日。右面板:1994年9月8日。(重绘Jeffrey et al。11。)
cpd水柱或海冰。Prezelin等。[88]证明了感应片冰的cpd藻类从南极附近接触后表面辐射4 h在早上。没有损伤诱导下- a +票面价值光合有效辐射)或票面价值。最近的一项研究在气候温和的巴伊亚法典(丘布特、阿根廷)面积证明光合抑制和DNA损伤形成pro -和真核远洋生物由于UYR [53]。在这个研究中,浮游生物样本在不同深度在水中孵化列,之后累积损伤测量的下午。cpd积累迅速在微生物群落保持表面,但在深度(3米和6米)减少观察,表明损伤被修复过程。清晰的深度依赖关系CPD丰富初夏被发现在南极海洋细菌和浮游植物组合,后者主要由硅藻[89]。在初夏,架子上层水的冰融化使得稳定层在这些沿海水域。在这里,细菌被发现含有更高水平的损害与真核生物相比分数(> 10 / mi)。然而,浮游植物表现出CPD丰度在整个夏天在这南极湾。夏季末降低太阳能的角度,降低水温和更高的意思风速造成水体成为混合。当时cpd仍出现在所有大小分数,但伤害是均匀分布在整个水柱。
固着生物没有下面的机会和破坏性的波长,因此可能拥有适当的策略来减少或防止DNA损伤。里昂等。[52]测量诱导的DNA损伤微生物群落与珊瑚粘液和观察到的CPD丰度随深度增加,一个明显异常的结果。DNA损伤在珊瑚粘液样本也始终低于生物采样在水柱类似的深处。这些结果表明,固着生物居住在较浅的水域更依赖光保护化合物如马斯河或利用更有效的修复系统。不幸的是,实验没有报道,可能会区分这些可能性。
上面描述的深度资料,要么是接近uv - b在中午达到顶峰,或在下午,CPD丰富假定为最大时,如果同意累计DNA有效的紫外线剂量。任何生物体的生理变化进行调查在白天期间,如修复系统的感应,可能阻止CPD积累模式后DNA有效剂量。因此,一昼夜的DNA损伤模式可以给最大CPD水平的信息,因为他们可能发生在白天还提供信息在原位修复或修复过程的感应。
Jeffrey et al。[11]表明DNA损伤积累白天bacterioplankton组合在墨西哥湾。他们发现,在日落之后立即CPD水平下降。在此期间的新DNA合成(测量胸苷掺入)没有高到足以解释cpd的cpd建议删除下降主要是由于切除(黑暗)修复和DNA损伤不是“稀释”增长[11]。此外,测量recA基因表达在这些样本[90]和bacterioplankton Gerlache海峡,南极洲(91、92),遵循一个明确的日常模式与最大表达日落之后。这些数据都表明,切除修复是必不可少的日常康复海洋bacterioplankton暴露于太阳。
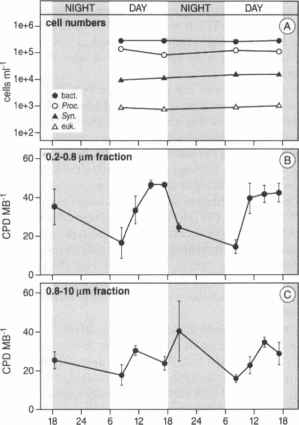
虽然有潜在的光致复活作用显著的证据在海洋bacterioplankton(91、93),进一步支持模型计算[85],其有效性原位的直接证据是有限的。Boelen等。[94]孵化CPD包含picoplankton样品在10米深度,生物有效表面uv - b含量只有6%的水平,但对紫外线A和PAR(参与photorepair)还高(见表1)。明显降低CPD水平没有发现,然而,在光周期。虽然不能完全排除,对紫外线a和PAR水平在10米深度不足以支持photorepair,光致复活作用似乎不流行的通路的CPD去除这些生物。此外,实验在亚喀巴湾,红海,还表示缺失或photorepair可以忽略不计,但主要是在夜间修复(图7)。Bacterioplankton孵化包在水面以及原位样品显示清楚日常模式与最大值的下午,虽然原位损伤水平较低的样本在[95](图7)。DNA损伤还发现白天积累在温带海洋细菌和浮游植物组合从巴伊亚法典,阿根廷。cpd积累迅速,当样本暴露于太阳辐射,即使是阴天[96]。> 10¡im分数(浮游植物,主要是硅藻)也accumu -
太阳时(h)
图7。一昼夜的CPD丰度在两个周期bacterioplankton大小分数在亚喀巴湾。(一)细胞主要浮游生物群体的数量;(B)原位采样在水面0.2 - -0.8 /即时通讯;(C)原位取样-10 / xm 0.8水面。(重新从
太阳时(h)
图7。一昼夜的CPD丰度在两个周期bacterioplankton大小分数在亚喀巴湾。(一)细胞主要浮游生物群体的数量;(B)原位采样在水面0.2 - -0.8 /即时通讯;(C)原位取样-10 / xm 0.8水面。(重新从
迟来的损失,尽管以低得多的利率比picoplankton分数。此外,这项工作还表明,光合抑制和CPD积累不同的日常模式。这个之前还发现热带淡水组合[97](的的喀喀湖,玻利维亚),当孵化整个表面的光周期。这里,CPD积累了整整一天,而光合抑制了uv - b几乎是常数。在巴伊亚法典孵化实验,阿根廷,没有发现修复当海洋微生物组合在photoreactivating光孵化。损伤累积时没有删除在下午时间在上午时间样本暴露于紫外线a - f PAR或独自不相上下。没有重大photorepair,因此,被认为有助于白天观察到的快速积累的cpd [96]。这些观察是按照类似的实验在南极(Buma未发表的结果)photorepair轻微或没有在uv - b的试验被排除在外。相比之下,原位修复被发现在巴伊亚法典在样本孵化3和6米水柱,uv - b的水平很低,但对紫外线a和PAR明显偏袒photorepair [53]。
Weinbauer et al。[12]显示一昼夜的cpd和6 - 4模式PPs在自然病毒社区地表水的墨西哥湾。白天一般CPD浓度增加,最高浓度被发现15:00h至18:00 h。第二天早上样本显示,损坏是在夜间。6 - 4页显示一个类似的趋势为cpd发现,除也显示在接下来的夜晚。然而,作者认为,这不仅减少可能是由于hostmediated修复也稀释的病毒复制的结果。在另一项研究中,威廉et al。[98]显示清晰的微分剂量反应photoreactivating和non-photoreactivating条件下病毒传染性。证明host-mediated修复是可以恢复很大一部分的传染性病毒,从而使病毒来完成他们的裂解周期。最近,原位测量表明,多达52%的太阳能radiation-inactivated病毒可能photoreactivated沿海海洋环境。
总之,所有的DNA损伤诱导的一昼夜的出版周期海洋bacterio浮游植物表明,即使photorepair发生时,它将发挥有限的作用。Photorepair并不妨碍快速建立白天的损害。因此,损伤积累模式经常发现DNA大致遵循有效的紫外线剂量,特别是在中午。
远洋鱼的卵和幼虫可能对uv - b诱导DNA损伤敏感,因为他们是小,透明和发生在海洋的上层。uv - b诱导CPD形成和修复能力进行了研究在新蛋和卵黄幼虫北部的凤尾鱼,Engraulis mor-dax,暴露在电离辐射[99],卵和幼虫死亡暴露在太阳辐照度。在较低的水平,即更自然的水柱的条件,有一个清晰的一昼夜的周期二聚物的浓度。这种模式密切关注太阳能强度(图8),而不是DNA有效剂量,为发现细菌和浮游植物(见上图)。这一昼夜的循环被认为是由于损伤和真正的photorepair的瞬时相互作用,而黑暗(切除)修复被证明是次要的。光致复活作用可能会停止当样品转移到黑暗。未孵化的胚胎,催生了在黑暗中,也表现出很强的photorepair响应,表明光裂合酶表达这些生物不依赖于先前的紫外线辐射机制。检查者等。[99]的结论是,CPD浓度时的取样是一个很好的指标的累积剂量和剂量率,而不是鳀鱼有一个高效photoenzymatic修复系统。协议,有效photorepair能力鳀鱼之前被描述[100]显示增加幼虫photoreactivating条件下生存。
很明显,需要更多的信息在条件影响photo-reactivation水生生物。首先,实地测量给contradic -
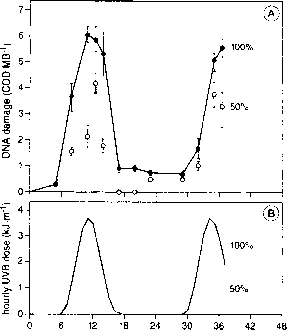
从午夜天时间(累计小时1)图8。北部的CPD浓度未孵化的胚胎鳀鱼:(A)新催生了鸡蛋,暴露于完整的太阳能辐照度(大胆的实线)或减少50% uv - b(轻实线)。(B)紫外线剂量率测量的意思是四个扫描(每小时达光电OM 752光谱仪),集成在280到320纳米之间。
(从检查者et al . 99年重绘。)
从午夜天时间(累计小时1)图8。北部的CPD浓度未孵化的胚胎鳀鱼:(A)新催生了鸡蛋,暴露于完整的太阳能辐照度(大胆的实线)或减少50% uv - b(轻实线)。(B)紫外线剂量率测量的意思是四个扫描(每小时达光电OM 752光谱仪),集成在280到320纳米之间。
(从检查者et al . 99年重绘。)
保守党结果photoreactiva-tion的重要性和有效性。同时,高效的光致复活作用潜力已经报道了大量各种各样的水生生物,当暴露在人工(光)条件。因此,即使存在潜在的光致复活作用,其他因素如营养和能源供应或人工灯和差异太阳光谱可能会干扰修复系统的感应。此外,紫外线的照射会导致水生生物的生存能力损失原位,从而关闭CPD移除。这种可能性诱导的DNA损伤修复的能力之外将在下一节中加以解决。
9.5.3残余DNA损伤和生存能力
几乎所有领域的研究已经开展到目前为止已经证明在日出前所谓的残余DNA损伤的存在。残余DNA损伤的发生表明,损伤诱导期间之前的uv - b照射事件并不是完全被各种修复途径之前,一个新的紫外线曝光周期(第二天)。
残余DNA损伤被发现在清晨样本Jeffrey et al。
[11]bacterioplankton从墨西哥湾和在南极海洋细菌和浮游植物[89]。CPD水平发现夏末在南极海域似乎支持的存在一个不可修理的分数:尽管发现了显著的残余CPD水平低入射紫外线水平和深入混合水列进一步减少意味着uv - b细胞经历。孵化实验还表明,cpd诱导在早上时间没有被photoreactivating光在下午(对紫外线a和紫外线b排除在外,承认,Buma,未发表的结果)或任何其他修复过程(即。,暗修复)。也发现类似的结果在温带海洋浮游生物组合从巴伊亚Bus-tamante,阿根廷[96]。事实上,残余DNA损伤报道每一个位置CPD丰度进行了研究。阿卡巴湾的实例(图7),picoplankton大小分数保留残余DNA损伤(14至43 CPD MB”1)结束的时候晚上尽管CPD的数量在黑暗中有所下降。这剩余损伤表明,黑暗无法删除所有cpd修复过程。典型Synechoc-cocus或细菌细胞含有大约2.1 x 10 ~ 15 g DNA细胞[101],这将意味着在50到160 cpd每个细胞仍然存在,阻止DNA复制和细胞分裂。Quaite等。[34]发现最初的二聚体频率较低(少于大约30 CPD MB-1)苜蓿幼苗没有使用切除修复。这个级别(30 CPD MB-1)匹配的残留水平发现早上亚喀巴海湾的样品。解释剩余损失水平的另一个建议是,DNA损伤并不是均匀分布在所有的细胞,但大部分的损伤积累几严重受损细胞,或变得无法修理。 These cells would then lose viability and eventually disappear by lysis [102].
实验室研究表明,紫外线照射导致海洋硅藻丧失生存能力[103 - 105]。事实上,缺乏维修报告在许多领域实验表明存在不可行的CPD包含细胞原位,有时进一步支持的光合作用同化率低,例如,巴伊亚法典区域,剩余CPD水平极高的[53]。一般来说,即使在人口受到相同的uv - b辐射剂量,DNA损伤并不是均匀地分布在细胞。Buma等。[10]表现出明显的不均匀分布的CPD硅藻细胞特定的荧光在人群中,从高标准差对CPD意味着特定的荧光,使用免疫化学的CPD标签结合流式细胞术(图9)。当推断这个领域,大多数的损害会积聚在有限数量的细胞。因为细胞分裂不能修复所有的损害之前完成,部分的人口可能会因此在正常方式复制,而最终受损的细胞溶解死亡和消失的。如果生存能力损失发挥着作用,残余CPD水平不仅取决于可行性损失率,但也不能存活细胞在水中的停留时间列。例如在阿根廷的海,剩余CPD水平很高,而CPD诱导率并不极端。这些高初始水平,因此,可能是由于在水中不能存活细胞的积累长期列。
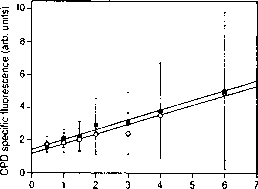
图9。剂量反应曲线CPD特定荧光对uv - b的剂量Cyclotella sp.细胞,用流式细胞仪测量。满圆:G1细胞,开放圈:G2细胞。误差线,标准差对G1和G2意味着细胞(至少4000个细胞分析/条件/细胞周期阶段)。(重新形成Buma et al。10。)
图9。剂量反应曲线CPD特定荧光对uv - b的剂量Cyclotella sp.细胞,用流式细胞仪测量。满圆:G1细胞,开放圈:G2细胞。误差线,标准差对G1和G2意味着细胞(至少4000个细胞分析/条件/细胞周期阶段)。(重新形成Buma et al。10。)
在这种情况下,这将是有趣的考虑潜在的营养/底物限制和uv - b压力之间的相互作用。营养的增加已报告减少bacterioplankton生产紫外线辐射的敏感性[91]。已经证明了[106]的很大一部分bacterioplankton在海洋水域是新陈代谢活动由于衬底的限制。浮游植物也可能经历养分(氮、磷、铁)限制在公海或post-bloom情况。次优的代谢活动在这些细胞将阻碍DNA修复,并因此导致快速积累这些细胞的DNA损伤。反过来,损伤积累会减少生存在这个社区的一部分。低维修率和残余DNA损伤(早上样品)结合低增长水平估计在许多bacterioplankton实地研究似乎支持这一假设。
9.5.4不同臭氧浓度的影响
到目前为止,很少有研究解决臭氧的影响损耗在海洋生物的DNA损伤积累。马洛伊等。[107]在南极ichtyoplankton跟着DNA损伤环境紫外线通量在南国之春。他们表明,自然水平的紫外线臭氧损耗造成重大的损害多细胞生物占领期间南极生态系统中营养水平较高。特别是,银鱼蛋被证明是容易CPD积累。此外,他们表明,CPD浓度银鱼蛋密切关注每日紫外线通量,表明损伤修复容易在一天内,按照发现的检查者et al .[99]鳀鱼蛋。
最近,Maedor等。[108]跟着每日浮游微生物的DNA损伤水平孵化环境下太阳能条件,包括臭氧波动,在帕默站,南极洲。虽然模式复杂,出现的变化之间的关系入射太阳辐射由于臭氧损耗和浮游生物的DNA损伤。Huot等。[85]开发了一个模型诱导的DNA损伤和修复在混合和非混血bacterio-plankton环境。同时,臭氧损耗的影响在混合和非混血系统整合。发现臭氧厚度对DNA损伤的积累造成最大的影响,与混合的影响相比,DOM浓度或叶绿素浓度[85],
继续阅读:维吉尼亚•维勒范创造出克里斯蒂娜Sundbck毛皮L菲格罗亚和E沃尔特·赫尔
这篇文章有用吗?