光保护机制
保护淡水水生生物从UV-induced损伤依赖于多种因素,可以作为光保护机制。紫外线辐射违反光保护机制足够时,UV-induced伤害发生。水生生物不同紫外线照射的宽容。有可能占据的生态位有机体之间的相互作用及其紫外敏感性。整个有机体的生命阶段,它的栖息地和习惯可能会补有机体对紫外线的忍耐。夜间或黄昏活动制度显然限制紫外线照射,将生物体UV-limiting栖息地的选择。鱼类已经适应了某种程度的紫外线和太阳紫外线辐射可能表现出不同的公差范围。自然物种适应高水平的太阳能辐射暴露容忍高紫外线水平将超过物种适应低水平的太阳能辐射。事实上,剃刀鲸吸盘,鱼类自然适应太阳能紫外线水平高,是模拟UV - b的宽容,没有开发晒伤后21天的接触[42]。相比之下,晒伤在彩虹鳟鱼和Lahontan切喉鳟两天内,和Apache鳟鱼(Onco-rhynchus Apache)后五天暴露于模拟uv - b [2]。
紫外线辐射的敏感性将太阳辐射随鱼的适应。提高水质和水化学的变化(特别是有机碳含量的减少)可以提高紫外线强度在水里列[43]。即使鱼可能检测紫外线应用在实验室的研究中,他们可能不直接避免太阳uv - b[44],但间接避免紫外线通过寻求职位低的水柱或寻求阴影避免激烈可见或紫外线a。因为太阳的可见光和紫外线a不会与臭氧损耗、增加更高水平的太阳能紫外线会被鱼和可能造成有害影响[45]。然而,鱼,利用表面或浅水栖息地可能进化适应容忍高uv辐照度的栖息地[46],
也有生理特点,从根本上提高有机体的宽容与紫外线包括photorepair机制受损DNA的效率。DNA是特别容易受到紫外线的,因为它引发cross-linkages的形成,或者在嘧啶基地胸腺嘧啶二聚体,。这些cross-linkages包括cyclobutane-type胸腺嘧啶二聚体、胞嘧啶,和尿嘧啶;嘧啶加合物;photohydrates;和DNA蛋白质使交联[47]会干扰DNA复制的细胞分裂和蛋白质合成必要增长和更换,可导致肿瘤的发展,以及病变。大多数生物能够修复DNA损伤诱导通过切除修复,uv - b光致复活作用,和复制后修复([47],见第9章),其中光致复活作用由DNA photo-lyase,推广一种酶,这种酶结合环丁烷二聚体,成为激活从紫外线a和可见光吸收光子,然后劈开前从环二聚体解开[48]。布劳斯汀等。[49]发现,这种酶在胚胎两栖动物的数量直接相关物种的uv - b的宽容。尽管这种相关性并不表明较高的二聚体修复在宽容的生物敏感生物相比,它确实表明细胞photorepair效率的基础。然而,研究结果与树蛙(Rana sylvatica)表示响应光裂合酶的变化取决于环境条件和史密斯等。[50]得出估算两栖动物photorepair是一个复杂的过程,之前的结论对于photorepair和两栖动物数量下降之间的关系必须评估。 Photorepair is likely to be ubiquitous among fish given the range of species for which evidence for photorepair has been found, including goldfish [51], anchovy larvae [52] and fathead minnow [53]. Photorepair efficiency in fish varied by as much as 500% between two closely related species [54].
切除修复包括损伤识别,切口附近的DNA链的DNA切除的病变和受损后的网站,和结扎后分离的DNA聚合酶[48]。物种可能有能力这两种类型的修复机制,可能会有所不同,哪一个占了主导地位。例如,大部分通过photorepair DNA修复发生在白天,和剩余修复发生在黑暗中通过切除修复[55]。
不管等photorepair有机体的效率机制并不完全有效,所以不要完全保证潜在的伤害。因此,适应更多的保护机制。这些包括色素隔离高活性氧自由基或其他活性物种产生的紫外线和负责DNA损伤和其他细胞损伤([56]参见第8章)。UV-shielding颜料还提供紫外线保护的一种手段。这些可以是外在以及内在的过滤器。水生动物提供光利用可用的环境因素,如医生,减少环境紫外辐射,因此生物接收剂量。水生生物也将防紫外线或反射颜料在他们的皮肤。
13.4.1光保护水生生物的物质
有几种类型的光保护水生生物中发现的物质(见第10章)。光保护色素有一定的一般特征如下面[57]。短暂,大多数光保护物质分享rc-electron系统发生在单双键交替等共轭键结构在线性分子,包含电子共振和芳环化合物。重叠的轨道7 r-electrons吸收最大值在紫外区域,导致一个精力充沛的7 r-electrons过渡到反键7我*电子轨道。改变结构的共轭分子的吸光度变化特征,因此减毒的发光光谱。长波长吸收较大的分子,吸收波长发生变化随着共轭数量的债券或取代基数量的增加。吸收增加更多的侧链和替换被添加到分子结构。非成键电子的氧、氮、和卤素原子成为环形结构的共振和转变的一部分吸收最大。在进化的过程中,开发了针对特定分子结构的吸光度的变化特征。有机体可以屏幕广泛的uv - b和a波长通过合成一系列photo-absorbing分子。
Mycosporine-like氨基酸,另一种类型的光保护物质,被发现在一个生物的多样性从细菌到鱼[57]。到19种氨基酸mycosporine-like已确定。某些生物含有这些物质的几个屏幕紫外线[58]。这些物质的浓度成比例地增加强度紫外辐照度他们是暴露于[58-60]。Mycosporine-like氨基酸被鱼类和无脊椎动物可能不是合成的,但通过饮食,尤其是从放牧藻类[61]。Gadusol,也认为是photo-protective,结构相关mycosporine-like氨基酸和发现鸡蛋的鳕鱼和Mediteranian在盐水虾鱼[62]和[63]。
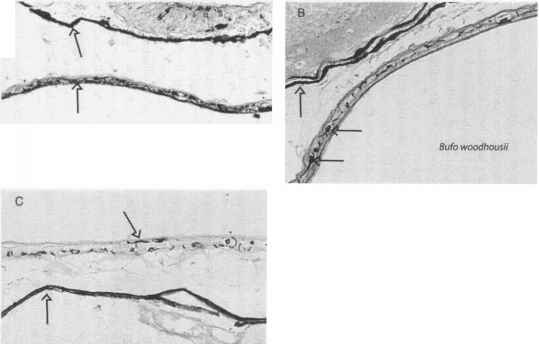
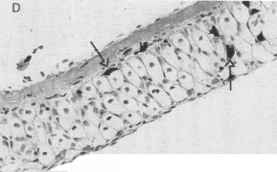
光保护黑色素被发现在脊椎动物和无脊椎动物生物的多样性和聚合物形成于5日、酚醛6-dihydroxyindole indolic复合[64]。黑色素是复杂的分子,广泛吸收紫外和可见辐射,但没有特定的吸收最大;然而,它们的吸收波长增加而减少(65、66)。在载黑素细胞产生黑色素,黑色素沉积在亚细胞细胞器称为黑素体,通常定位在细胞核[67]。暴露于紫外线会导致黑色素的增加生产和黑素体的数量在长期暴露在紫外线[57]。鱼一般都没有离开这个皮肤表皮层中的黑色素相对紫外线辐射的保护,和似乎依赖于一种无色化合物(s)分泌的粘液覆盖身体提供表皮光[68]。在青蛙(图4),然而,在表皮黑色素发生,包括光似乎与黑色素的数量及其分布[69]。UV-B-tolerant北方蟾蜍(以北风)有明显的双层的黑色素。相关柴棚的蟾蜍(以woodhousii)生活在低海拔的栖息地也有一个双黑色素层,但是,似乎黑色素分散和集中。夜间灰树蛙(雨蛙癣)一层黑色素,并对紫外线很敏感。火蜥蜴(钝口螈属tigrinum)有扩散和有限的黑色素细胞的分布和高度敏感的uv - b(凯瑞个人沟通)。
类胡萝卜素色素广泛发生在甲壳类浮游动物,尽管成分和数量随物种[55]。类胡萝卜素是由藻类、细菌和植物和无脊椎动物和脊椎动物通过食物链转移[70]。类胡萝卜素被认为是主要的photoabsorbing桡足类色素。Copopods显示的类胡萝卜素浓度大于水蚤类的,和那些生活在紫外线强烈的环境浓度高于生物从低紫外线栖息地[71]。卡罗-
以北风之神
雨蛙杂色的
图4。幼虫皮肤的黑色素分布(A)北方蟾蜍(以北风之神),(B)柴棚的蟾蜍(以woodhousii), (C)灰树蛙(雨蛙癣),(D)老虎蝾螈(钝口螈属tigrinum)。注意致密层的黑色素在皮肤(单箭头)的蟾蜍和青蛙幼虫和黑色素分布在表皮层(双箭头),这是北方蟾蜍紫外线最密集的宽容,和最密集的夜间(和紫外线敏感)灰树蛙。黑色素分布老虎蝾螈幼体是分散在整个真皮和表皮(箭头)和密集的青蛙和蟾蜍相比少得多。[(A, B)修改小et al。[69];(D)从凯里未发表的数据。)
^ _钝口螈属tigrinum
以北风该组织
雨蛙杂色的
图4。幼虫皮肤的黑色素分布(A)北方蟾蜍(以北风之神),(B)柴棚的蟾蜍(以woodhousii), (C)灰树蛙(雨蛙癣),(D)老虎蝾螈(钝口螈属tigrinum)。注意致密层的黑色素在皮肤(单箭头)的蟾蜍和青蛙幼虫和黑色素分布在表皮层(双箭头),这是北方蟾蜍紫外线最密集的宽容,和最密集的夜间(和紫外线敏感)灰树蛙。黑色素分布老虎蝾螈幼体是分散在整个真皮和表皮(箭头)和密集的青蛙和蟾蜍相比少得多。[(A, B)修改小et al。[69];(D)从凯里未发表的数据。)
^ _钝口螈属tigrinum tenoids往往分布在整个身体的桡足类,而枝角目动物倾向于集中在卵巢脂质类胡萝卜素和鸡蛋[55]。除了类胡萝卜素,黑色素也存在于角质层水蚤类的,往往是更为明显的物种生活在高海拔地区(72、73)。类胡萝卜素紫外线空气净化能力有限,然而,在水蚤类的类胡萝卜素被认为发挥重要的光保护作用,隔绝氧自由基(55岁,56)。
淡水鱼随uv - b[2]的宽容。宽容不亲热的接触模拟uv - b似乎无关的黑色素的色素沉着,但与一位身份不明的无色的光保护皮肤的物质[74]。这光保护物质似乎局部外背侧皮肤层(表1),其中包括表皮和上覆粘液,但也发现的眼睛,腮UV-tolerant剃刀鲸吸盘(Xyrauchen texanus) [68]。下面的例子说明了这个光保护物质的数量在一个特定的鱼类物种的紫外线耐受有关。通道鲶鱼被发现对模拟紫外线非常敏感,黑暗的曝光和24小时内没有检测到光保护物质[17]。剃刀鲸吸盘接触太阳能模拟uv - b没有出现晒伤后21天的接触,而在48 h[68]虹鳟鱼晒伤。当部分背皮肤未被鱼和甲醇提取扫描分光光度计,有更多的光保护物质的提取razor-back吸盘比彩虹鳟鱼[42]。因此,有直接关系的光保护物质,这些鱼的时间开发了晒伤(表2)。光保护物质似乎是一个自然防晒霜和保护剃刀鲸吸盘从模拟太阳紫外线的有害影响。黑色素形成鲜明对比,这提供了一些光在任何给定的波长,光保护
组织 |
物种 |
LN) 1 |
Amountb |
外背侧皮肤层 |
虹鳟鱼 |
294.0 (0.0) |
0.10 (0.0) |
剃刀鲸吸盘 |
294.5 (0.2) |
0.44 c [0.05] |
|
内心的背侧皮肤层 |
虹鳟鱼 |
290.2 (0.4) |
0.03 (0.01) |
剃刀鲸吸盘 |
292.3 (0.6) |
0.12 c [0.01] |
|
的眼睛 |
虹鳟鱼 |
没有检测到 |
- - - - - - |
剃刀鲸吸盘 |
293.5 (0.3) |
0.02 (0.003) |
|
鳃 |
虹鳟鱼 |
没有检测到 |
- - - - - - |
剃刀鲸吸盘 |
293.5 (0.3) |
0.02 (0.002) |
|
肝 |
虹鳟鱼 |
没有检测到 |
- - - - - - |
剃刀鲸吸盘 |
没有检测到 |
aMean Amaj(标准误差)六个鱼。
bMean吸光度单位/毫克湿重(au / wt)(标准误差)的组织六个鱼。cPhotoprotective因素之间的组织明显不同的物种。
aMean Amaj(标准误差)六个鱼。
bMean吸光度单位/毫克湿重(au / wt)(标准误差)的组织六个鱼。cPhotoprotective因素之间的组织明显不同的物种。
表2。相对多的防晒霜在鱼皮,平均紫外线照射诱导晒伤,和鱼类紫外线伤害的脆弱性;编制小和Fabacher[2],尤因等。[17],Fabacher和小(68、74)
天的脆弱性暴露U V
物种数量的防晒霜晒伤损伤
剃刀鲸抽油100 > 21低
色素青鳉59 10中
白化青鳉59 10中
虹鳟鱼23 2高
通道鲶鱼ND 1非常高
波长(nm)
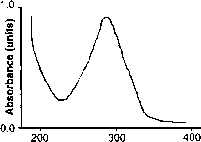
图5。光谱光度测量的光保护物质的紫外吸光度的皮肤剃刀鲸抽油(Xyrauchen texanus)。吸光度最高出现在294 nm,吸光度曲线的宽阔的肩膀会提供保护,免受紫外线波长小于和大于294海里。(从Fabacher和小68。)
波长(nm)
图5。光谱光度测量的光保护物质的紫外吸光度的皮肤剃刀鲸抽油(Xyrauchen texanus)。吸光度最高出现在294 nm,吸光度曲线的宽阔的肩膀会提供保护,免受紫外线波长小于和大于294海里。(从Fabacher和小68。)
物质将提供大量的保护紫外线的波长因为吸收最大范围在294 nm和斜率的肩膀覆盖许多紫外线波长峰值(图5)。
当切喉鳟和剃刀鲸吸盘被暴露于模拟uv - b,切喉鳟雄clarki henshawi)显示严重可见迹象暴露(背侧皮肤变暗)48 h[15],剃刀鲸吸盘,然而,没有任何可见的晒伤的迹象在整个72 h的接触。切喉鳟大大减少的光保护物质比剃刀鲸吸盘。切喉鳟背侧皮肤的组织学检查显示粘液细胞脱落,坏死和水肿在表皮和真皮,而且,在某些情况下,继发真菌感染(图6)。相反,组织学观察剃刀鲸抽油的皮肤显示坏死发生,但严重脱落,坏死中观察到切喉鳟皮肤没有发生(图7)。有剃刀鲸吸盘表皮厚度的增加,显然造成肥大和增生的大型细胞含有大量低电子密度的中部地区。这些
继续阅读:Tevini斑鳟属
这篇文章有用吗?

