辐照度的影响在北极水域的生产力和养分供应对气候变化的看法raybet雷竞技最新
Jean-Eric Tremblay和乔纳森·盖格农
部门生物魁北克海洋,Pavillon Alexandre-Vachon,大学拉瓦尔,魁北克,QC,加拿大祝福OA6,(电子邮件保护)
文摘
先前公布的数据分析表明,每年远洋北冰洋线性相关的初级生产的持续时间无冰的时期,可能通过累积暴露在太阳辐照度。然而,无冰时间最长的地区位于周边海域和冰穴,养分供应由平流或风和对流引起的垂直混合可以广泛。随后补充营养的强烈驱使初级生产中未达到的水平分层内部(如波弗特海),除了上升流区域。公布数据的再分析显示累计生产之间没有关系入射太阳辐射在生长季节。我们建议年单位面积上的初级生产季节性变化无冰水域主要由控制环境迫使氮供应。偶然变化光政权应该主要影响时间和可能的物种组成主要生产脉冲(s)上混合层,底下,浮游植物的利用营养低透光层。而持续增加的热量和淡水供应北冰洋应该加强垂直分层和进一步阻碍意味着向上的营养供应,情景还直接上层海洋的大气强迫可能与长期暴露在光和协同作用大大增强远洋的生产力。
介绍
北冰洋的物理环境变化迅速,深刻(适配器2005)。最引人注目的表现是最低程度的下降9月海冰变薄的多年冰
(郭et al . 2007;马斯兰尼克et al . 2007;斯特罗伊夫et al . 2008年)。冰盖变得本质上是季节性的,在一些地区,融化越来越早或形式越来越晚(Lemke et al . 2007年)。河量在上升,增加淡水内容和有机和加载无机物在沿海地区(彼得森et al . 2006年)。极端风事件的强度和频率增加了在一些地区(Zhang et al . 2004年),这将影响上层海洋动力学下reduced-ice场景。
定量和预测海洋初级生产这些变化的响应是一个先决条件理解他们对北极的影响食物链,生物地球化学循环和海气通量的主动式气候气体。raybet雷竞技最新一个明显的持续变化的结果是,到达海洋表面的光量增加。不那么明显的是这种能源补贴与伴随的扰动的水柱来改变大小和远洋初级生产的物种组成。自养生长需要光,但也需要营养物质在海洋上的库存是有限的。理想的混合营养生长变化的比率在浮游植物类群和不同营养成分各不相同,在源水的弥漫北冰洋(Tremblay et al . 2002 b)。一小部分本地回收这些营养素的生物活性(即排泄、分解),但整体生产力的变化必须靠外来的氮(N)补贴的透光层。这个输入可能源自深海硝酸,大气中的氮气或降水和河流的无机或有机N。
在现在的环境下的快速变化,值得问的概括可以从数据聚集在过去50年里帮助支架对未来的预测,并提供一个视角。初级生产的几个评论发表对俄罗斯海域(Vetrov和Romankevich 2004),北极(例如Sakshaug 2004;勒让德et al . 1992;史密斯和Sakshaug 1990)和冰穴(Arrigo 2007;Tremblay史密斯,2007)。这些评审文档北极海洋生态系统的多样性和复杂性,不同的地区冰动力学和物理海洋学不同广泛(见麦克和Wassmann 2006)。
为了合成pan-Arctic数据,Rysgaard et al。(1999)获得之间的正相关年度初级生产和无冰的持续时间。表面上,这种关系表明,初级生产将在未来,增加成正比的延长无冰的季节。我们评估是否Rysgaard et al .(1999)提出的相关调查因果有预测价值,潜在因素。我们首先考虑分别辐照度和营养的作用,总结了一些观点和假设,在未来的研究。
数据挖掘
我们回顾文献考虑了研究Rysgaard et al .(1999),使用Sakshaug(2004)和Vetrov Romankevich(2004)除了最近的工作在巴芬湾及波弗特海(克莱因et al . 2002;辛普森et al . 2008年)。合成Rysgaard et al .(1999)公布的年度报告价值的初级生产但没有上下文所需的物理和化学参数控制因素的深入分析。后者需要信息的深度测量法采样,养分供应和日期的第一个和最后一个季节性测量初级生产(需要估计事件辐照度测量期间)。
虽然本身的信息,一些研究认为被排除在分析之外,因为他们被认为不具有可比性。抽样的网站底部深度和盐度小于5 m和5%啊,分别(霍纳和施克拉德1982;好的et al . 1993;Meskus 1976)属于浅海,潮间带或河流生态系统和被解雇。模型结果也折扣(例如Slagstad和Stole-Hansen 1991;田口1972;Wassman et al . 2006年)一样不能被视为在同一基础实际测量分析。
两个级别的数据合成。一级只关注那些研究填充时间序列(即测量在频繁的时间间隔在一个受限区域或水质量)的初级生产。时间序列,并没有解决主要生产脉冲的峰值和尾巴结束(格兰杰1980 - 1968;沃尔什et al . 1989;亚历山大1974;麦克罗伊和戈林1976;尼尔森和汉森1999)被排除在一级数据基础。为了简明、清晰、比较、课程报告的时间和比较累积主要生产(EP)获得的价值超越连续时间点之间的集成。2级给年度初级生产价值估计插值和/或推断,最好是由作者自己或我们最初计算时没有完成。在某些情况下,这样做是通过将数据从2年或以上。 We assume that the level of confidence attached to the different estimates is much higher for level 1 than for level 2 since extrapolations do not take into account possible changes in the time courses when measurements stopped long before the ice cover was re-established. The number of studies considered for level 1 (9 studies and 13 distinct time courses) is lower than the number of studies considered for level 2 (n = 12).
在大多数研究中,数据在事件或水下太阳辐射是不提供用于比较的目的。每日平均事件短波辐射(W m - 2)纠正云层从NCEP / NCAR再分析检索。对于每一个时间序列,计算累积辐射在海面乘以每天平均在瓦特每平方米86400年代和总结这些每日剂量超过60天或全职课程。
除非另有规定,结果表示在GJ(千兆焦耳)m - 2。信息初始浓度的营养生长季节前缺席一些研究。在这些情况下,浓度被从区域或前或后研究从2005年世界海洋数据库(Boyer et al . 2006年)。
数据集的一般性质
一般用于研究水平1和2的位置分析是图1所示,每个采样点的特征表1中给出。研究网站集群在西方北极,一个意想不到的地区偏见,和范围广泛的纬度(60 - 82.5°N)深度(15 - 450,作为对于一个给定的平均采样地区)和年日测量初级生产(从3月下旬到7月上旬)。
-
 and annual estimates (all symbols) of primary production. Numbers refer to the studies described in Table 1.")
- 图1所示。采样地点的地图用于详细的分析时间序列(黑色圆圈)和年度初级生产的估计(符号)。数字参考表1中描述的研究。
 and annual estimates (all symbols) of primary production. Numbers refer to the studies described in Table 1.")
ID年 |
位置 |
经度纬度深度(°N) (°W或°E) (m) |
开始结束(一天)(一天) |
参考 |
|||
1 |
1959年 |
迪斯科湾(w .格陵兰岛) |
69.250 |
53.567°30 W |
129年 |
141年 |
彼得森于1964年 |
1 b |
1960年 |
迪斯科湾(w .格陵兰岛) |
69.250 |
53.567°W |
93年 |
165年 |
彼得森于1964年 |
2 |
1959年 |
哑铃湾海(林肯) |
82.517 |
62.167°W 25 |
172年 |
66年 |
Apollonio 1980 |
3 |
1968年 |
白令海 |
60.000 |
170.000°W 100 |
- - - - - - |
- - - - - - |
麦克罗伊和戈林1976 |
4 |
1968年 |
n格陵兰岛 |
82.167 |
31.233°E 26 |
- - - - - - |
- - - - - - |
安徒生1981年 |
5 |
1968年 |
汉湾 |
63.667 |
68.450°W 50 |
176年 |
110年 |
遍布全球1980 |
5 b |
1969年 |
汉湾 |
63.667 |
68.450°W |
167年 |
82年 |
遍布全球1980 |
6 |
1971年 |
坚决湾 |
74.684 |
94.868°W 15 |
181年 |
102年 |
韦尔奇和Kalff 1975 |
7一个 |
1973年 |
迪斯科湾(w .格陵兰岛) |
69.167 |
53.500°W 50 |
171年 |
119年 |
安徒生1977年 |
7 b |
1974年 |
迪斯科湾(w .格陵兰岛) |
69.167 |
53.500°W |
138年 |
168年 |
安徒生1977年 |
7 c |
1975年 |
迪斯科湾(w .格陵兰岛) |
69.167 |
53.500°W |
81年 |
77年 |
安徒生1977年 |
8 |
1976年 |
波弗特海 |
71.000 |
148.000°30 W |
- - - - - - |
- - - - - - |
1978年凯莉 |
8 b |
1977年 |
波弗特海 |
71.000 |
148.000°W |
- - - - - - |
- - - - - - |
1978年凯莉 |
8 c |
1976年 |
Chuckchi海 |
71.800 |
156.000°W 40 |
- - - - - - |
- - - - - - |
1978年凯莉 |
8 d |
1977年 |
Chuckchi海 |
71.800 |
156.000°W |
- - - - - - |
- - - - - - |
1978年凯莉 |
9 |
1982年 |
巴伦支海 |
76.000 |
32.500°E 200 |
98年 |
181年 |
雷伊et al . 1987 |
10 |
1996年 |
年轻的声音(如格陵兰岛) |
74.310 |
20.251°35 W |
174年 |
One hundred. |
Rysgaard et al . 1999 |
11 |
1998年 |
不。巴芬湾 |
76.000 |
74.000°W 450 |
98年 |
104年 |
克莱恩et al . 2002 |
12 |
1993年 |
不。格陵兰海 |
80.230 |
13.000°W 200 |
- - - - - - |
- - - - - - |
克莱恩et al . 2002 |
13 |
2004年 |
SE。波弗特海 |
70.807 |
125.393°W |
152年 |
84年 |
辛普森在新闻等人 |
虽然第一次测量不同的日期之间的广泛研究,不同的时间课程(要求等级2)策划了一个共同的起源时间为了直接比较的影响时间EP(图2)。一个让人震惊的结果是,在大多数生产系统EP达成渐近线而在别人通常大约线性增加。后者观察挑战一般认为北极系统强烈脉冲在时间和生产高峰期持续几个星期。图2的另一个值得注意的方面是,虽然增加的初始利率不同,课程迅速承担最后的时候,相对等级。这样分层意味着没有一次连续体EP值躺上打开水期的持续时间增加。换句话说,这些系统的初级生产水平在很大程度上是初条件或一组和EP的总体差异不解释为打开水期的持续时间本身。多达85%的变异性EP结束点是可预测的只有60天后,尽管测量时间持续3到8个月。
50个25
0 30 60 90 120 150 180天运行以来的第一次测量
图2所示。时间的累积初级生产(EP),开始的第一天测量每个时间序列。不同的符号引用ID数字在表1中给出的不同的研究和抽样年:1(开环),1 b(露天广场),2(开放的钻石),5(开放六边形),5 b(开放的三角形),6(打开倒三角形),7(灰色圆),7 b(灰色的平方),7 c(灰色钻石),9(灰色六角)10(灰色三角形),11(黑色圆圈),13(黑色广场)。
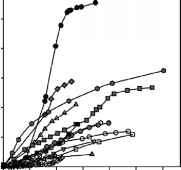
0 30 60 90 120 150 180天运行以来的第一次测量
图2所示。时间的累积初级生产(EP),开始的第一天测量每个时间序列。不同的符号引用ID数字在表1中给出的不同的研究和抽样年:1(开环),1 b(露天广场),2(开放的钻石),5(开放六边形),5 b(开放的三角形),6(打开倒三角形),7(灰色圆),7 b(灰色的平方),7 c(灰色钻石),9(灰色六角)10(灰色三角形),11(黑色圆圈),13(黑色广场)。
辐照度的作用
冰盖和极端的太阳活动周期实施严厉限制初级生产。这些限制是最严重的钢管,消失多年冰与极夜,使光合作用不可能通在大部分的冬季。条件不太严格的冰在低纬度地区变量动态和辐照度政权为初级生产者创造一系列短暂的利基市场。更换多年冰的季节性冰可能会创建一个更有利的光在高纬度地区的气候。raybet雷竞技最新光在这方面扮演着两种角色。它决定当净浮游植物的增长是可能的和累积能量是多少用于光合作用过程中生长的季节。
一旦光已达到水浮游植物的可用性是由多种因素决定的,包括混合、集平均辐照度被细胞和影响水透明度通过沉积物re-suspension浅货架上(例如森林et al . 2008年),浮游植物的生物量和水平投入的溶解和颗粒有机质在沿海地区。事件辐照度之间的相互作用、水的透明度和混合深度确定潜在的浮游植物的增长经验的考虑,不会重复的主题在这里(Nelson和史密斯1991;Sakshaug和Slagstad 1991)。值得注意的是我们的分析中使用的数据集很少包括所需的信息估计平均辐照度的混合层。
开始生产的季节
净初级生产认为可能只有当浮游植物有足够的光为正增长,一般按照斯维德鲁普斯的临界深度模型修正和Passow (1990)。尽管社区位于冰水界面或顶部的透光层可能不是大力混合,他们需要足够的光线来超过损失,即辐照度必须高于补偿光强度低于浮游植物呼吸作用的总和和其他损失(放牧、溶解和下沉)超过获得的碳通过光合作用。最近的经验估计表明,补偿辐照度的硅藻社区在北极的意思类似于北大西洋(1.3 - -1.9摩尔广达m - 2兴奋不已;Tremblay et al . 2006 b)。
-
 the year-day of the first measurement, and (B) the latitude of the sampling site.")
- 图3所示。之间的关系的意思是,在海平面下降短波辐射测量和(A)的第一个星期期间year-day第一测量,和(B)采样站点的纬度。
 the year-day of the first measurement, and (B) the latitude of the sampling site.")
不足为奇的是,开放的位置在年初获得更少的辐照度最初(图3)。意思是事件短波辐射测量表面在第一周的独立于纬度(图3 b),表明生产开始时控制本质上是由区域海洋动力学过程和冰而不是太阳周期。一旦有足够的光净增长发展的初始速率布鲁姆应该取决于暴露的微藻收获这光辐照度和能力有效,由细胞的生理状态的影响,可能温度(Tremblay et al . 2006 b)。
累积辐照度
图4所示的模式是反直觉的。系统与初级生产的最高水平后60天(相比,所有的系统都可以)收到更少比最高效的光。这个结果符合硅藻无法收获的光突然从附近切换完成后立即默默无闻快速冰消融后满阳光(Tremblay et al . 2006 b),一个可能是最严重的问题在系统营业到很晚,接近夏至。
藻类如何应对光在生长季节的发病可能取决于快速发生转变,而且在不同的photoadaptation和photoacclimation战略种子的生物礼物(Sakshaug和Slagstad 1991)。的光抑制浮游藻类的有时被认为是一个实验性的人工制品的孵化项目,即细胞是在高亮度超过他们做下混合,原位条件。快速的出现,非侵入性的活性荧光技术表明,光抑制往往是严重的在这些社区,即使暴露在光闪烁的几微秒时间内。事实上,这种短期急性光抑制通常比明显从孵化项目跨越几小时到一天(原位或模拟现场),这表明在后者,藻类有时间调整和恢复,在即时测量信息的不干预。
正如预期的那样,辐照度明显负面影响在考虑时间课程全部消失。然而,辐射的总量(EE)收到期间课程与EP没有关系,要么期间每一次课程(图4 b)或每年(图4 c)。还请注意在图4 b, EP的EE值的数量级范围1.5 - -2.1 GJ m - 2。虽然严格的比较基于混合层中的辐照度是不可能回想起来,很明显,不同的辐照度剂量是目前不足以占EP跨seasonally-open系统的范围。
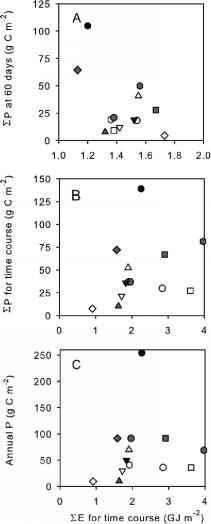
图4所示。累积之间的关系主要生产(EP)和累计下降,海面(EE)的短波辐射(A)的前60天每一次课程,每次课程的总持续时间(B),和(C) 1年EP与EE计算时间进程(B)。符号按图2所示。
图4所示。累积之间的关系主要生产(EP)和累计下降,海面(EE)的短波辐射(A)的前60天每一次课程,每次课程的总持续时间(B),和(C) 1年EP与EE计算时间进程(B)。符号按图2所示。
营养物质的作用
移置硝酸
营养物质提供给上层北冰洋的各种流程操作在不同的时间和空间尺度。水平提供输入河流和电流来自邻近海域。立即平流尤为重要下游太平洋和大西洋的网关,它可以支持强烈的初级生产,尤其是在楚科奇海。地表水驻留在中央北极ca。10年,除非他们的交通发生只在多年冰,不稳定的营养成分很容易消耗在外围的第一年。这种情况也适用于无机营养物质由主要河流。例如,马更些河供应大量的硅酸盐北冰洋,但是任何残留,无机磷酸盐和氮前耗尽淡水羽进步到海里(Emmerton et al . 2008;辛普森et al . 2008年)。
脱氮的浅架子亚北极太平洋和北极,结合高硅酸加载河流确保磷和硅留在过剩,特别是在太平洋海域占主导地位。氮是主要的产量限制营养在北极筋疲力尽首先在水华研究到目前为止(Kattner和Budeus 1997;辛普森et al . 2008;Tremblay et al . 2002 b, 2008),甚至在巴伦支海的硅酸盐和磷酸盐浓度相对较低(Reigstad et al . 2002年)。
物理奇点偶然给定地区的营养补贴。这些奇异点可以采取内波的形式,风暴侵蚀盐跃层,shelf-break上升流和动态不稳定引起的深度测量法或对流(马西斯et al . 2007;威廉姆斯et al . 2006;Zhang et al . 2004;Tremblay et al . 2002年)。上升流事件的发生率和力量和盐跃层扰动可能增加的频率和强度上升气旋(杨et al . 2004年)和常年的撤退冰袋在架子上打破(麦克和查普曼2003)。目前还不可能的评估或预测的最终结果的变化意味着与情景交付的营养级和初级生产的物种组成。
初级生产者的总可用性营养是初始库存之和的透光层(冬季)和任何输入发生一旦赛季开始增长。因为这些补贴是难以量化,很少被认为在报纸上用于级别1和2的分析,我们只计算的初始库存营养出席或发病前生长季节。
很明显的深度分布研究(表1),纵向一体化必须标准化比较。获得的时间序列在深水表明明显季节性营养赤字经常扩展到50 - 75米(Tremblay et al . 2002;史密斯et al . 1997年),由于情景在生长季节的垂直混合或浮游植物生长在地下的能力叶绿素最大值(Tremblay et al . 2008年)。在这个视图中,不到50米底深处的系统不能得到相同的新产品,因为他们有限的养分之间的垂直距离或重叠库存和沉积物反硝化和透光层。在年轻的声音,为例,浅底(35米)和相关的脱氮限制硝酸盐浓度2 | m在冬天春天(总库存70更易与m - 2) (Rysgaard et al . 2004年)。不到40米深度水域浮游植物硝酸通常获得初始库存150±135更易与m - 2,而在更深的水域可以利用1000更易与m - 2(图5)。为了执行一个合法比较浮游植物的养分可以,晚库存估计上层75 m /深水浅或整个水柱网站超过75米。
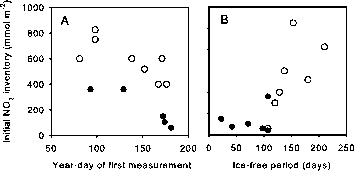
图5所示。硝酸的初始浓度之间的关系出现的生长季节和(A)的year-day级别1和2的第一个测量数据,和(B)估计持续时间的无冰期,所有数据。封闭和开放的圆圈表示底深处少或超过30 m,分别。
图5所示。硝酸的初始浓度之间的关系出现的生长季节和(A)的year-day级别1和2的第一个测量数据,和(B)估计持续时间的无冰期,所有数据。封闭和开放的圆圈表示底深处少或超过30 m,分别。
硝酸虽然最浅的水域有最小的库存,深度测量法仅解释了30% (p < . 05)整体可变性的初始硝酸盐的可用性,表明在深地点物理过程起决定性作用。为例,系统位于大西洋北极的部门(如北水和巴伦支海东部,即研究9和11个表1)通常是得天独厚的表层水的更大的敏感性相对于强烈的垂直混合分层Pacific-derived水域(例如阿蒙森湾,研究13日在表1)(参见Tremblay et al . 2008年)。总的来说,初始硝酸盐库存显示负相关关系(r2 = 36, p < . 05)的year-day第一测量(图5)和积极的(r2 = .41点,p < . 01)的估计时间无冰季节(图5 b)。这种模式意味着系统表层海水长时间暴露于大气中体验一个更大的氮负荷。这种效应的可能原因是,系统与长期的无冰期,如北方
水或巴伦支海暴露在冬末或秋季和对流风暴侵蚀疲软的垂直分层和补充营养(Tremblay et al . 2002年)。春末夏初开放的水域(如东北水)当风较弱和太阳诱导——融化水分层强看到每年的垂直补给营养(Kattner和Budeus 1997)。风暴的巴伦支海躺在铁轨在夏天也经历显著的营养更新haline分层适中(Sakshaug和Slagstad 1992;Wassman et al . 2006年)。在极端的情况下,即使是强大的分层描述加拿大盆地的水可以被推翻,从而提供充足的营养,否则贫困上透光层。这个条件是观察到的巴芬湾西北部,但潜在的混合过程尚未证实(Tremblay et al . 2002 b)。
硝酸的初始库存上75被证明是相当可靠的预测的EP在60天(r2 = 53, p < . 01;图6 a),每次课程结束时(r2 = i, p <措施;图6 b)。这些关系解释为什么年产量可以从累积值预测60天后,即初级生产的总体水平在很大程度上是预先确定的初始养分负荷虽然光可能影响的初始利率下降。硝酸加载还举行年度P之间的关系,但这一次北水显然站在正异常。去除离群值的分析产生一个非常健壮的关系(r2 =尾数就p <。;图6 c)。异常高水平的年度初级生产在北方水混合是由情景,布鲁姆和生产力的持续下降至10月(qr et al . 2007;克莱恩et al . 2002年),这表明硝酸的初始库存只提供有限的氮供应系统。这种限制的另一个例子是发现在一些地区的白令海,持续营养更新导致不同寻常的生产率水平类似于和高于观察到在北方水(例如Sambrotto et al . 1984; Springer et al. 1996). Clearly, the inshore portions of shallow Arctic shelves do not experience a significant late-season, vertical re-supply of nitrate simply because the deep reservoir is absent. At the shelf break or in deeper regions, episodic mixing and upwelling may augment primary productivity far beyond the levels expected from initial conditions. It follows that, on a regional basis, the effects of climate change on primary productivity will greatly depend on bathymetry and distance from the shelf break, rivers and the adjacent Pacific and Atlantic oceans.
多元回归模型,其中包括硝酸EE除了库存仅略提高EP的预测,和部分系数对辐照度的影响不显著(没有显示)。显然在这种背景下,氮负荷是主要的控制汽车列车的生产力。考虑到一些研究温度数据是可用的,我们无法找到一个温度和EP的初始增加之间的关系(参见Tremblay et al . 2006 b)。
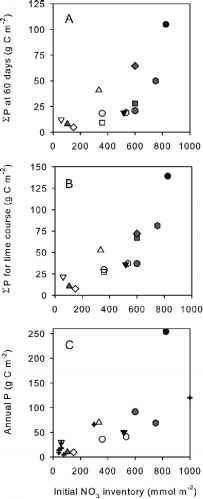
图6所示。初始浓度的硝酸发病之间的关系(A)的生长季节和EP每次课程的前60天,(B)的总持续时间每一次课程,和(C) 1年。符号按图2 C,除了面板crossesrepresent研究的年度估计但没有时间课程(ID 3、4、8个模拟和12在表1)。
图6所示。初始浓度的硝酸发病之间的关系(A)的生长季节和EP每次课程的前60天,(B)的总持续时间每一次课程,和(C) 1年。符号按图2 C,除了面板crossesrepresent研究的年度估计但没有时间课程(ID 3、4、8个模拟和12在表1)。
新的和再生生产的贡献
移置N吸收由自养生物(即新产品)最终用于浪费或损失由运输或下沉。的一些废物留在透光层可以直接重用或容易分解成有用的N形式由光化学或细菌。N因此回收硬盘再生生产,导致总初级生产但不能维持净收益的增加有机质,生态系统生产力和垂直碳出口(Dugdale称和戈林1967;埃普利和彼得森1979)。的相对重要性和再生生产必须解决理解驱动pan-Arctic总产量的差异和产生这些变化意味着什么收获的资源和海气通量的主动式气候气体。raybet雷竞技最新
随着无冰季节的持续时间增加,有一个明显的可能性,EP也只是因为N周期多次透光层。调查这种可能性是有用的考虑出版/比率的估计,即移置N吸收比总N吸收。理想情况下,所有来源的吸收外来的和应该考虑回收N /比率,但只有的净吸收硝酸盐、铵,有时,尿素通常估计。
可比性,填充时间序列/比例的解决的一个主要部分生长季节仍然是极其罕见的。这个数据是对北方水(/比率= 0.58;Tremblay et al . 2006年)和东北水(/比率= 0.65;史密斯et al . 1997年),重复测量了类似的一段ca。3个月开始在春天。这些/比率非常相似,尽管三倍硝酸不同初始库存和四倍的区别在两个系统之间的年产量(252 g C m - 2北和东北66 g C m - 2水)。比较表明,再生生产占总初级生产的很大份额,但仅仅是成正比的新的生产和供应的移置联合国两个系统的特点是可以忽略不计的库存以来铵和尿素在生长季节的开始,一个转换/比率,(1 - /)/ /可以用来估计的次数N最初在硝酸回收三个月期间,收益率低0.6 - -0.7的值。在北方水、净量920更易与N m - 2是消耗4月23日和7月1日之间(Tremblay et al . 2006年)。到那时,只剩下100更易与N m - 2彩球和更易与N m - 2铵+尿素的透光层(Tremblay et al . 2002 a, 2006 a)。这种粗糙的预算意味着高达87%的硝酸N最初接受的透光层居住在不到68天,这是相对于无冰的持续时间周期短(ca。6个月)。在这个视图中,长期无冰的季节应该促进再生生产只要有一个持续的或外部N来维持它的脉冲电源。
影响,视角和未来的研究
评估当前和未来的生产力的北冰洋应该区分率和产量限制过程,和新的和再生之间的生产。光和浮游植物的生理状态影响营养物质消耗的速度(Tremblay et al . 2006 b)早期和晚期(qr et al . 2007年)年,而外部氮的供应上屈服的透光层集有机物生产(Tremblay et al . 2006 b)。离岸,多年冰的消融将首先增加单位面积上的新的生产剩余养分徘徊,但随后几年的刺激减少,除非新的物理机制促进复发性营养更新通过垂直混合,上升流或涡流创世纪。在浅货架库存有限的无机N和一个世俗的,季节性的冰盖,怀疑旷日持久的无冰期,将增加新的生产单位面积从架子上休息,河流和邻近的海洋。seasonally-open水从本质上讲,这更大程度上应该增加盆地规模初级生产,但超出了最初从多年过渡到季节性冰,不能在和自己导致持续,数量级增加单位面积上的EP(由Rysgaard et al . 1999年)在目前的地区。
汽车列车的工作环境变化很可能改变N的透光层的上行供应。这些变化是否导致补贴或一个给定地区的贫困将很大程度上取决于淡水平衡的变化与大气上层海洋的强迫。而变暖,降水和淡水的增加交付应该进一步提高垂直分层和减少营养物质的通量向上在沿海地区,蒸发和出口多年冰离岸有利于盐渍化et al。(2007),通过弱化分层的表层可以使它更容易混合的风暴。这种敏感性也增加了冰雪覆盖的收缩时期,而提交地表水直接风迫使在早春或深秋(例如Tremblay et al . 2002年)。
平流的作用和河流直接、水平的营养来源仍然是难以评估。历史上,河流没有提供大量的无机氮和磷北冰洋,但放电和贡献的冻土融化富裕水域可能增加营养负荷在沿海水域(弗雷et al . 2007;麦克勒兰德et al . 2007年)。这种补贴,然而,可能是本地以来自然无机营养物质很容易消耗近海。溶解有机氮的场景可能不同(不)逃早期消费和潜在支撑在沿海水域初级生产的一部分。
东南波弗特海最近的数据显示,无机磷和溶解无机碳的净损耗(DIC)硝酸后继续疲惫(辛普森et al . 2008;Tremblay et al . 2008年),这表明自养生物获得一个备用源的移置联合国的一些河流提供的可以直接使用一些浮游植物或可用的光化学或细菌攻击non-readily不稳定化合物(例如Vahatalo和泽普2005)。新的证据表明溶解有机质的输入和生物可用性在春季洪水比先前认为的基础上夏天的数据(福尔摩斯et al . 2008年)。过量的磷和低硝酸盐的可用性高北极也使得它的沃土N2固定。Yamamoto-Kawai et al。(2006)表明,过量的磷维持下游N2固定在北大西洋,但它也可能是在北极的,特别是和在条件(如温度)是diazotrophs有利。这个假设实验还有待证实,但它似乎是合理的,区域N2固定将获得的重要性随着上层北极变暖,会更加分明。
大部分的文学处理河道流量可以理解关注溶解有机碳(DOC)输入和它们对海洋生物地球化学的影响,包括异养和光化学释放DIC(例如Belanger et al . 2006年)。二氧化碳的这个版本对海气通量的影响,然而,可以减轻附带增加造成的净初级生产N的放松限制。这是有可能的,然而,水透明度下降可能引发从营养限制转向光限制在浅架子。对这些方面的综合研究除了N2固定,硝化作用和沉积物反硝化作用是考虑到他们的潜在影响的空气——保证海交换二氧化碳和一氧化二氮(例如首领et al . 2008年)。
正在进行的物理环境的变化可能会影响分类的优势在生产脉冲(例如Arrigo et al . 1999;沃尔什et al . 2004),但这个话题仍严重可以理解在北极。除了周边海域(即白令海峡,巴伦支海,拉布拉多和格陵兰岛),莉亚娜huxleii或殖民地棕囊藻属pouchetii但是此属可以达到较高的生物量中,记录了浮游植物水华在北冰洋的内部主要由硅藻。没有令人满意的解释提出了这个地理隔离,但在南大洋,不同的光适应政权(Arrigo et al . 2007年)可能发挥作用。还需要更多的研究在这个问题上,从分类地位的变化对食物链产生深远影响,生物地球化学通量和气候反馈(例如Tozzi et al . 2004年)。raybet雷竞技最新
另一个未知的是如何减少复发的时间季节性冰盖和冰藻的生产将影响sympagic-pelagic-benthic交互。从水体中藻类可以被困在新的冰或殖民的底部厚一年级冰在冬季和春季,但反过来明显要少得多。一方面,有证据表明,冰藻类,释放冰时,立即水槽底部和底栖生物提供食物社区,尤其是在浅大陆架(勒让德et al . 1992年)。另一方面,保留腐冰藻类在上层水体中已经观察到(Michel et al . 1993年)和推断为远洋食草动物提供重要能源碳预算的基础上(Michel et al . 1996年)。程度ice-grown藻类目前远洋种子和形状是一个开放的问题。虽然不影响的整体收益率有机物质在生长季节,时机和物种组成的改变远洋华可能会改变食物网和食草动物的成功更灵活的生活史。
总之,本文表明之间的正相关关系的持续时间无冰期和年度初级生产Rysgaard et al .(1999)观察到没有预测能力本身。这种关系本质上是由深度测量法和微分易感性不同的海洋省份垂直混合。浅的生产力或强烈分层北极水域不会赶上弱分层或upwelling-prone地区因为生长季节的持续时间增加。这样当然会增加调节年度生产率在给定的水域,尤其是地下通过增加生产叶绿素最大值,但变化不会在EP达到数量级范围内观察到pan-arctic规模。几乎所有的短期响应是可能的在这个信封,最近Arrigo et al .(2008)所示为2007年的生长季节。然而,持续增加单位面积上的初级生产只会发生较大的地表水每年太阳辐射相匹配的反复出现的营养补贴。是否这发生本质上取决于相伴的净结果淡水平衡的变化,营养水平加载和大气强迫上北冰洋。
确认
我们感谢丹尼·杜蒙特及时帮助与NCEP再分析太阳辐射数据。这项工作是飞机的赠款支持加拿大自然科学和工程研究理事会和ArcticNet卓越中心的网络,并对项目的贡献的海洋魁北克和加拿大研究主席北极海洋生态系统对气候变化的响应。raybet雷竞技最新
引用
适配器(2005)科学报告:北极气候影响评估。raybet雷竞技最新剑桥大学出版社、剑桥
亚历山大·V(1974)初级生产力近岸波弗特海的政权,参照冰生物群的潜在作用。:里德JC,萨特我(eds)。波弗特海的海岸和货架,北美北极研究所,卡尔加里安徒生OGN(1977)初级生产,照明和Jorgen Bronlund水文学
峡湾,北格陵兰岛。Medd om Granland 205:1-27安徒生OGN(1981)的年度周期浮游植物的初级生产和水文学在迪斯科海湾地区,西格陵兰。Medd om Granland 6:3 - 65 Apollonio年代(1980)初级生产在北冰洋的哑铃湾。3月杂志61:41-51 Arrigo KR(2007)的物理控制初级生产在北极和南极冰穴。:理发师DG,史密斯WOJ (eds)。冰穴:Windows世界,爱思唯尔,海洋系列,阿姆斯特丹
沃森写DL Arrigo KR,罗宾逊DH,邓巴RB, DiTullio GR, VanWoert M, Lizotte议员(1999)浮游植物群落结构和营养物质的减少,二氧化碳在南大洋。科学283:365 - 367 Belanger年代,谢HX Krotkov N, Larouche P,文森特•WF Babin M (2006) Photomineralization北极海域陆源溶解有机物的从1979年到2003年,气候变化和年际变化的影响。raybet雷竞技最新全球Biogeochem周期。doi: 10.1029/2006 gb002708凯里AG)、小(1978)。海洋生物。:环境评估阿拉斯加大陆架临时合成波弗特/ Chuckchi, NOAA,博尔德CT
卡马克E,查普曼直流(2003)风力货架/在北极大陆架盆地交换:冰盖的联合作用范围和shelf-break深度测量法。“Res。gl017526 doi: 10.1029/2003
卡马克E, Wassmann P(2006)食物网和physical-biological耦合pan-Arctic货架上:统一概念和comprehensice视角。学监Oceanogr 71:446 - 477首领RA, LaRoche J, K阿尔提耶里,Arrigo KR,贝克AR,卡彭DG,康奈尔大学的年代,Dentener F,加洛韦J, Ganeshram RS, Geider RJ, Jickells T, kuyper MM, Langlois R,丽丝PS,刘SM,船帆JJ,摩尔厘米,Nickovic年代,Oschlies,皮德森T,普洛斯彼罗J, schlitz R, Seitzinger年代,索伦森噢,Uematsu M, Ulloa O,沃斯M,病房B,萨莫拉L(2008)在开放海域大气人为氮的影响。科学320:893 - 897 Dugdale称RC,戈林JJ(1967)的初级生产力的新和再生形式的氮。Limnol Oceanogr12:196 - 206 Emmerton CA, Lesack LFW,文森特WF (2008) Mackenzie河营养交付的北冰洋和效果麦肯齐三角洲在开放水域条件。全球Biogeochem周期doi: 10.1029/2006 gb002856埃普利RW,彼得森BJ(1979)颗粒有机物质通量和深海浮游新的生产。自然282:677 - 680森林,Sampei M, Makabe R,佐佐木H,理发师D•布兰顿Y, Wassmann P L福捷(2008)的年度周期颗粒有机碳出口在富兰克林湾(加拿大北极):环境控制和食物网的影响。JC004262 J地球物理学研究》doi: 10.1029/2007
弗雷克麦克勒兰德JW,福尔摩斯RM,史密斯LC(2007)气候变暖的影响和冻土融化在河边的喀拉海运输氮和磷。raybet雷竞技最新J地球物理学研究》doi: 10.1029/2006我,jg000369 qr Gosselin M,克莱因B, Tremblay我Fouilland E(2007)新的和再生生产在夏末盛开一个北极冰湖。Mar Ecol-Prog Ser 345:13-26格兰杰嗯(1980)的佛洛比西尔湾的初级生产,北极的加拿大。:邓巴乔丹(ed)。
海洋生产机制,剑桥大学出版社、剑桥福尔摩斯RM,麦克勒兰德JW,雷蒙德PA,弗雷泽BB,彼得森BJ,施蒂格利茨M(2008)易变性的DOC经由阿拉斯加北冰洋的河流。“Res。doi: 10.1029/2007 gl032837霍纳R,施克拉德GC(1982)的相对贡献冰藻类、浮游植物、底栖微藻在近岸地区初级生产的波弗特海。北极35:485 P - 503把裙边,Alasaarela E,宽松的HG, Jokela年代,Storgaard-Envall C(1993)初级生产的季节性变化和营养浓度的沿海水域Bothnian湾和夸克。Aqua芬Kattner 23:165 - 176 G, Budeus G(1997)东北水冰湖的营养状况。J 3月系统10:185197
克莱恩B,勒布朗B、梅ZP贝雷帽R, Michaud J,曼迪CJ,冯Quillfeldt CH,加纳我,罗伊,格拉顿Y,科克伦JK, Belanger年代,Larouche P, Pakulski JD, et al .(2002)浮游植物生物量、产量和潜在的出口在北方水。深海Res II 49:4983 - 5002
郭R,坎宁安GF, Zwally HJ易D (2007)冰,云,土地高度卫星(卫星)北极海冰:检索的干舷。112年J“Res: jc003978 doi: 10.1029/2006
勒让德L, Ackley科幻,Dieckmann GS, Gulliksen B,霍纳R, Hoshiai T, Melnikov IA, Reeburgh WS,斯宾德勒米,沙利文连续波(1992)生态学的海冰生物群。2。全球意义。极地生物Lemke 12:429 - 444 P,任J,小巷RB,艾莉森,我卡拉斯科J,弗拉托G,藤井裕久Y,卡瑟G, Mote P,托马斯•RH张T (2007)。观察:变化,雪,冰以及冻土。所罗门:et al。(eds)。raybet雷竞技最新2007年气候变化:物理科学基础工作组的贡献我的第四次评估报告政府间气候变化专门委员会,剑桥大学出版社,剑桥,英国/纽约
施特略夫马斯兰尼克J,福勒C J, Drobot年代,Zwally HJ, D,金刚砂WJ(2007)一个更年轻、更薄北极冰盖:增加潜力快速广泛的海冰损失。“Res。doi: 10.1029 / 2007 / GL032043马修斯特里,Pickart RS, Hansell哒,Kadko D,贝茨NR(2007)涡运输楚科奇货架:有机碳和养分的影响上盐跃层西北冰洋。J地球物理学研究》doi: 10.1029/2006 jc003899麦克勒兰德JW,施蒂格利茨M,潘F,福尔摩斯RM,彼得森BJ(2007)最近的变化在硝酸和溶解有机碳Kuparuk上游的出口,美国的阿拉斯加的北坡。J“Res-Biogeosci。doi: 10.1029/2006 jg000371麦克罗伊CP,戈林JJ(1976)年度预算的主要生产在白令海。3月Sci Commun 2:255 - 267
Meskus E(1976)的主要生产东北Bothnian湾。《大学奥卢42:55 - 62
米歇尔·C,勒让德L,塞JC, Demers年代,范德维德T(1993年)春天冰藻和浮游植物组合之间的耦合东南部哈德逊湾,加拿大的北极。极地生物13:441米歇尔- 449 C,勒让德L,英格拉姆RG, Gosselin M, Levasseur M(1996)碳预算的海冰藻类在春天:一个重要的证据转移到浮游动物食草动物。J“Res 101:18345 - 18360
纳尔逊DM,史密斯我们(1991)斯维德鲁普回顾-临界深度,最大的叶绿素水平,和控制的南部海洋生产力irradiance-mixing政权。Limnol Oceanogr 36:1650 - 1661尼尔森TG,汉森BW(1999)浮游生物群落结构和碳循环在格陵兰岛的西海岸夏季分层情况。1。水文学、浮游植物和bacterioplankton。Aquat活细胞生态16:205 - 221彼得森GH水文学(1964),主要生产、水深、和“Tagsaq”迪斯科
海湾,西格陵兰。麦克勒兰德J Medd om Granland 159:1-45彼得森BJ,咖喱R,福尔摩斯RM,沃尔什我Aagaard K(2006)的变化轨迹北极和亚北极淡水循环。科学313:1061 - 1066 Polyakov IV、Alexeev VA Belchansky GI, Dmitrenko IA,伊万诺夫VV, Kirillov SA Korablev AA,斯蒂尔M, Timokhow洛杉矶,Yashayaev我(2007)北冰洋淡水在过去的100年里变化及其原因。气候raybet雷竞技最新21:364 J - 384 Reigstad M, Wassmann P,立管连续波,Oeygarden年代,雷伊F(2002)水文学的变化,营养和叶绿素a的边际冰区和中央巴伦支海。J 3月系统38:1-2
雷伊F, Skjoldal人力资源,Slagstad D(1987)初级生产与气候变化在巴伦支海。:Loeng H (ed)海洋环境的影响商业鱼类资源的分布和种群动态在巴伦支海,学报第二Soviet-Norwegian研讨会,海洋研究所,卑尔根Rysgaard年代,尼尔森TG,汉森BW(1999)季节性变化在营养,远洋初级生产和放牧在北极高纬度地区的沿海海洋生态系统,年轻的声音,格陵兰岛东北部。3月生态掠夺Ser 179:13-25 Rysgaard年代,Glud RN, Risgaard-Petersen N, Dalsgaard T(2004)脱氮氨氧化活动在北极海洋沉积物。Limnol Oceanogr 49:1493 - 1502 Sakshaug E(2004)主要和次要生产北极的海洋。:斯坦R,麦克唐纳
RW (eds)。有机碳循环在北冰洋,施普林格,柏林Sakshaug E, Slagstad D(1991)光和极地海洋生态系统中浮游植物的生产力——一个生理的观点。极地Res 10:69 - 85 E Sakshaug Slagstad D(1992)海冰和风能:在巴伦支海对初级生产力的影响
海中。大气压海洋30:579 - 591 Sambrotto RN,戈林JJ,麦克罗伊CP(1984)的大型年度生产西方白令海峡的浮游植物。科学225:1147 - 1150
辛普森K, Tremblay J-E,格拉顿Y,价格海里(2008)年度研究养分分布的波弗特海东南部,Mackenzie架子和阿蒙森海湾。J地球物理学研究》doi: 10.1029/2007 JC004462 Slagstad D, St0le-Hansen K(1991)浮游生物生长动力学在巴伦支海:模型研究。极地Res 10:173 - 186。斯V, Passow U(1990年)春天开花启动和斯维德鲁普的临界深度模型。
史密斯Limnol Oceanogr 35:228 - 234 W, Jr .) Gosselin M,勒让德L,华莱士D,戴利K, Kattner G(1997)新产品在东北水冰湖:1993。J 3月系统10:199 - 209 Springer,麦克罗伊CP,弗林特MV(1996)白令海绿带:崖边生产过程和生态系统。施特略夫鱼Oceanogr 5:205 - 223 J, Sereeze M, Drobot年代,Gearheard年代,荷兰MM,马斯兰尼克J, Meier T, Scambos助教(2008)2007年北极海冰范围骤降。EOS事务。是地球物理学联盟89:13-14
田口年代(1972)数学分析的初级生产在夏季白令海。:Takenouchi (ed)生物海洋学的北太平洋北部,Idemitsu Shoten,东京
Tozzi年代,阿斯科菲尔德,法尔PG(2004)历史气候变化和海洋湍流作为两个关键的浮游植物官能团选择性raybet雷竞技最新药物。3月生态掠夺Ser 274:123132
加拿大我,史密斯WOJ (2007)。初级生产和营养动力学在冰穴。:理发师DG,史密斯WOJ (eds)。冰穴:Windows世界,爱思唯尔,海洋系列,阿姆斯特丹
格拉顿Tremblay我,Y, Fauchot J,价格海里(2002)气候和海洋强迫的,净和硅藻生产北水冰湖。格拉顿深海Res II 49:4927 - 4946 Tremblay我,Y,卡马克EC,佩恩CD,价格海里(2002 b)大规模北极环流的影响,北方水冰湖在巴芬湾营养库存。J地球物理学研究》doi: 10.1029/2000 jc000595 Tremblay我,服部年宏H,米歇尔•C Ringuette M,梅Z-P,洛夫乔伊C,福捷L,霍布森KA, Amiel D,科克伦JK(2006)营养结构和通路的生物碳流在东部北水冰湖。学监Oceanogr 71:402 - 425 Tremblay我,米歇尔·C,霍布森KA, Gosselin毫克,价格海里(2006 b)开花动态尽快开展北冰洋水域。Limnol Oceanogr 51:900 - 912 Tremblay我,辛普森K,米勒L,马丁•J•布兰顿Y,理发师D,价格海里(2008)垂直稳定,一年一度的营养物质和叶绿素荧光动力学在沿海,波弗特海东南部。J地球物理学研究》doi: 10.1029/2007 jc004547 Vahatalo AV,泽普R(2005)光化学溶解有机氮的矿化铵在波罗的海。环境科学工艺39:6985 - 6992 Vetrov AA, Romankevich EA(2004)俄罗斯的北极海域的生物产量。:Vetrov AA, Romankevich EA (eds)。俄罗斯北极地区海洋碳循环,施普林格,纽约。
沃尔什JJ,麦克罗伊CP,马车夫路,戈林JJ, Nihoul JJ, Whitledge TE,布莱克本,帕克PL, Wirick CD, Shuert PG, Grebmeier JM,施普林格,特里普RD, Hansell哒,Djenidi年代,Deleersnijder E、K•亨利,隆德英航,安徒生P, Mullerkarger铁、碳和院长K (1989)氮循环在白令海峡楚科奇海源地区organic-mjatter影响AOU北冰洋的要求。学监Oceanogr 22:277 - 359沃尔什JJ, Dieterle哒,Maslowski W, Whitledge TE(2004)年代际变化的生物物理迫使北极海洋食物网:数值结果。jc001945 J地球物理学研究》doi: 10.1029/2003
Wassman P, Slagstad D, Wexels立管C, Reigstad M(2006)造型巴伦支海的生态系统动力学包括边际冰区二世。碳通量和年际变化。J 3月系统59:1-24
韦尔奇他Kalff J(1975)坚决湾海洋新陈代谢,西北地区。环极北部生态会议学报》上。第二部分,NRC,渥太华Yamamoto-Kawai M,卡马克E, F麦克劳克林(2006)氮平衡和北极通流。Nautre 443:43
杨J, Comiso J,沃尔什D, Krishfield R, Honjo年代(2004年),猛烈的混合和对北冰洋的潜在影响。J地球物理学研究》doi: 10.1029/2001 jc001248张X,沃尔什我,张J, Bhatt, Ikeda M(2004)极地气旋活动的年际变化,1948 - 2002。气候raybet雷竞技最新17:2300 J - 2317
参考书目
辛普森公斤,Tremblay我,价格在阿蒙森海里(2009)营养动力学海湾和巴瑟斯特角冰湖:1。新产品在春天推断从营养提取资金。3月生态掠夺Ser(新闻)
史密斯我,Jr .)(1995)初级生产力和新产品在东北水(格陵兰岛)在1992年夏天冰湖。J“Res 100:4357 - 4370
继续阅读:在北极海域浮游植物的生产及其响应最近变暖
这篇文章有用吗?