农业集约化
农业集约化的概念描述了农业生态系统结构、功能、管理实践和目的在从“传统”到“现代”形式演变和变化过程中的变化。与我们目前关注的问题最直接相关的变化是种植的植物和牲畜物种越来越专业化(图11.1)。在现代主义思想中,专业化与生产效率的提高有关,尽管这在科学上是有问题的。例如,Rappaport(1971)指出,多物种家庭花园在能量上比单一栽培更有效。特定丰富度的下降通常伴随着同一地区土地利用率的增加和管理干预的加强,从而对劳动力和自然资源造成更大的压力。然后,这些资源被从外部购买的工业原料,如化肥、农药和石化能源所取代。这导致了对农业生态系统功能的内部(主要是生物)控制向外部(主要是经济)调控的转变。
因此,这些农业发展的后期阶段是通过与市场的联系而得以实现的,而且往往伴随着国家或区域经济其他部门的大量补贴。这往往会混淆的第二个主张现代农业在经济上,它比以自然资源为基础的系统更有利可图。
虽然所有这些因素都是农业集约化的一部分,但它们不一定同时发生,而且已经证明很难对各种各样的农业集约化进行分类农业生态系统分成几个容易描述的类别。尽管如此,基于土地使用和管理干预强度的松散分组为讨论生物多样性和生态系统功能之间的关系提供了一个有用的框架。以下是沿着这一梯度的一系列不同农业生态系统类型的简要描述,包括对其相对生物多样性的评论。这最容易用植物(特别是作物)和牲畜物种的多样性来描述,但区分这种计划的生物多样性和当地生物多样性是有用的,后者包括所有相关的但无经济生产力的植物(例如杂草、覆盖物种、农林业树木等)、动物(包括害虫和土壤动物)和微生物(例如病原体、共生体和微生物)土壤微生物).
11.2.2农业系统的多样性
几个世纪以来,热带三大洲的大片森林一直是各种形式的“转移农业”(也被称为“刀耕火种农业”)的所在地,当地也有各种各样的名称。这种广泛的农业形式的基本原则是短的作物阶段与长时期的自然或改良的休耕植被交替。因此,收益是在长期基础上管理的,而不是在短期内最大化(Ruthenberg 1980;Ramakrishnan 1992)。传统上,轮作农业系统通过混合种植系统在种植阶段保持多样性。在这个范围内,经济上重要的作物主要是一年生植物,多年生灌木和乔木被及时分离,并限制在森林的休耕再生阶段。根据农业周期和有关社区的社会和经济背景,混合作物种类的数量可能有很大差异,从6种到40多种不等。这种土地使用系统还可以包括家禽和猪等动物组成部分。作物阶段本身并不能表达系统的总体多样性;包括休耕期在内,植物物种丰富度可达数百种。
轮作休耕土地压力的增加或劳动力短缺可能导致刀耕火种农业形式的加剧。休耕期的缩短和久坐现象的增加通常导致同一块土地3-10年轮作一次。这种系统的休耕期通常以草本“杂草”物种为主,在土地准备耕作之前,这些物种可能被砍倒并在土壤表层分解。这一阶段的多样性因此大大降低。种植强度可能因地而异,从几种粮食作物到高度多样化的系统。豆科作物通常是混合种植制度的一部分,另外还有传统的谷物,如水稻或玉米,以及不太为人所知的粮食、药用和其他价值作物。
家庭花园是人类最古老的传统方式之一潮湿的热带地区在农业实践中模仿自然是通过将树木和其他多年生植物作为精心建造的家庭花园的组成部分,这是世界上许多地区普遍存在的系统。家庭花园是小块土地,通常0.5-2公顷,位于靠近居住地和施肥家庭废物.家庭花园有丰富的植物物种多样性(30-100种),以木本多年生植物为主,结构分层(Gliessman 1989;Nair 1989;Ramakrishnan 1992),混合了不同习性的一年生和多年生植物——草本植物、灌木、乔木和藤本植物——以及更传统的粮食作物。农民获得食物,柴火,药用植物香料和观赏植物,还有一些全年的现金收入。更复杂的是,家庭花园与传统的畜牧业系统,如家禽和猪的共同联系。这些自我维持的系统在生态和经济上都非常有效,但正在迅速消失。与轮作一样,这些系统的生物多样性总量与许多自然系统相当。
化合物的农场传统的农业潮湿热带地区的系统通常由同一家庭控制下的多种土地使用系统组成。子系统的范围可以从家庭花园到轮作休耕地,再到完全静止和相对专业化的田地(见下文)。西非(Okigbo和Greenland 1976年)以及亚洲(Ramakrishnan 1992年)都描述过这种复合农场。这些系统既具有空间复杂性,又具有较高的生物多样性。
耕地-牲畜混合耕作虽然轮作种植是潮湿热带地区的特征,但在干燥的热带草原地区,农业传统上以牲畜生产为中心。在非洲的许多地区,耕牛的人直接或间接地(通过产品的易货)从牲畜中获得几乎所有的需求,而定居的耕耘者则轮流或轮流地种植谷物(如高粱)和根茎作物(如木薯),两者之间存在着强烈的文化分离。
在大多数地区,随着政治边界的改变和土地压力的增加,游牧牲畜管理已经下降,这种分离已经被牲畜和粮食作物生产紧密结合的更固定的系统所取代。这种联系既是经济的,也是生态的。牛为耕地生产提供了耕田和运输的动力,也是施肥的粪肥来源。在放牧期间从牧场收获养分,并转移和集中在作物田里(Swift et al. 1989;Campbell et al. 1996)。这就需要对牛进行持续的管理,对农田进行间断性的集约管理,而且通常是男女分开的。这些系统的土地利用范围很复杂,在生产中使用的物种种类繁多,特别是来自放牧子系统的物种。
在上述所有系统中,树木都是常见的组成部分,要么是选择性地从自然植被中保留下来,要么是故意种植的。在世界许多地方,这一特点已得到进一步发展,以产生更有目的性的“农林业系统”。根据这些系统的结构和功能属性,已经确定了三大类(Nair 1989):农业是在同一块土地上使用作物和树木,包括灌木或藤蔓;森林畜牧业是牧草和树木的结合;农业畜牧业结合了粮食作物、牲畜牧场和树木。现代研究利用这些系统产生了一系列集约化的农林业实践,其中最著名的是“巷子耕作”。树木被紧密地成行种植,形成树篱,作物生长在它们之间的“小巷”中。因此,除了定期修剪树木以提供护根物或饲料并减少与作物的地上竞争影响之外,它与间作制度类似。因此,这些活动需要额外的劳动力,这是决定系统接受度的关键因素(Kang et al. 1990)。在这类系统中使用的树木通常因其多用途性质而被选择,但在实践中,集约化农林业的发展导致系统中使用的树木种质资源显著缩小。
间作和轮作土地利用的集约化最终可能导致一种或多种粮食作物的连续种植。这些实践在使用的多样性和复杂性方面有所不同。间作是指在同一土地上长期种植多种作物的做法。种数最多可达10-15种,但通常较少(2-6种)。尽管如此,就潜在产品的范围而言,这对农民来说仍然是一种分散风险的投资生态策略它可以拥抱(弗朗西斯1986)。
根据种植方法、农民的需要和用途,可以成行种植、随机分散种植或沿田地边缘种植。谷物和豆类的组合作为这种系统的“核心”很常见(例如,在半干旱的印度,高粱和豌豆,或珍珠小米和落花生),但谷物和谷物或谷物根茎作物的组合在西非也很常见。
轮作是一种强度相似的做法,即在不同的时间在同一块土地上种植不同的作物,即在连续的种植季节。轮作是世界许多地区集约化农业的一个共同特征,主要是因为需要避免疾病的累积。在谷物轮作中纳入豆科作物被公认为是提高肥力的一种手段,但在许多热带作物制度中很少保持严格的轮作顺序。
专业化经济作物系统集约化的一个常见结果是专业化农田比例的增加,其中一些专门用于“高价值”作物。它们通常构成传统经济的一部分,生产出可以用其他材料交换的产品。随着更结构化市场的发展,这些作物可能成为现金经济的重要组成部分。传统的经济作物包括各种果树、香蕉、生姜、菠萝、山药和特殊产品,如扫帚草(用于制作扫帚)或竹子(用于各种用途)。随着工业革命的到来,橡胶、可可、油棕或咖啡等小规模种植园作物被纳入其中农业系统在世界上许多地方。这些农田往往需要在一年中的某些时间进行密集管理,包括田间准备、病虫害管理、收获或收获后活动。尽管如此,他们传统上是内部处理的,劳动力完全来自家庭内部。其中许多系统还继续强调回收有机残基,对无机残基的依赖最小化肥.特殊作物通常与其他植物穿插在一起。在传统系统中,这些田地或种植园通常是非常多样化的(例如。东南亚的“丛林”橡胶),但现代种植园对其他物种的保护要“干净得多”(见11.2.4节)。
在世界许多地区,粮食作物,特别是谷物的领域专业化是现代(即二战后)农业的典型特征。这些系统代表了生物多样性的最终减少,即单一作物的基因统一、连续种植。这种形式的农业依赖于机械化耕作(汽油驱动),作物管理和收获。土壤和病虫害的管理受到化学调控,从而对微生物和无脊椎动物的生物多样性产生影响地上和地下(图11.1)。这种集约化水平包括热带地区的集约化水果种植园、加利福尼亚和地中海地区的集约化果园系统、全世界的集约化谷物生产以及大规模蔬菜生产等系统。
11.2.3农业集约化对生物多样性的影响
农业生态系统大致可以根据其管理强度来排序。广义梯度可能从不受管理的植被(通常是森林或草地)到“随意”管理(包括轮作、家庭花园和游牧牧业),再到低强度管理(包括传统的复合农场、轮作休耕和稀树草原混合耕作),再到中等强度管理(包括园艺、牧场混合耕作和巷子耕作),再到高强度管理(包括作物轮作、复种、巷作和间作),最后发展到现代主义(种植园、果园、集约化谷物和蔬菜生产)。
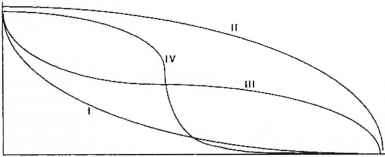
人们普遍认为,随着生境从森林到传统农业再到现代农业的变化,生物多样性会减少(Altieri 1990;Hoiloway和Stork 1991;Pimentel et ai 1992)。如果我们沿着强度梯度绘制总的生物多样性与点的关系,那么得到的关系很可能是单调和递减的。然而,曲线的确切形式是不确定的,图11.2中给出了四种可能的关系场景。在考虑假设曲线的形状之前,我们应该注意到x轴上强化水平的具体定位是近似的和非定量的。显然,要更严格地探讨这些关系,就必须推导出一个或若干个强化的定量指数。
曲线I假设,一旦人类的使用和管理对生态系统产生影响,生物多样性就会出现重大损失,这可能是对这种关系最普遍的预测。另一个极端。相比之下,曲线II的多样性只在高强度下受到显著影响,一般被认为不太可能。尽管如此,我们还不知道有任何数据或理论论据可以为在这两个极端之间做出决定提供确凿的证据。事实上,关于这个问题的少量数据(见下文)可能被解释为支持第二类曲线。然而,我们建议,在大多数情况下,介于这两种状态之间的东西将是模式。这种关系的两种中间形式由曲线III和曲线IV表示。
曲线III是生态学家预期的“温和”版本,它只是说,在生物多样性最初急剧减少之后,随着管理的加强,进一步的损失相对较小,直到达到真正现代系统的极限。曲线IV可能是一个更有趣的假设。
无管理系统(森林、草地)
休闲管理
(大栽培。
游牧畜牧业,
低强度管理(传统的复合农场,轮休,传统的
中等强度经营(园艺、牧场混合(武装、传统经济种植)
高管理(轮作、多套作、套作、间作)
无管理系统(森林、草地)
休闲管理
(大栽培。
游牧畜牧业,
低强度管理(传统复合农场、轮作休耕、传统家庭花园)农林复合
中等强度经营(园艺、牧场混合(武装、传统经济种植)
高管理(轮作、多套作、套作、间作)
现代主义(种植园和果园,密集的谷物和蔬菜生产!
图11.2农业集约化与农业生态系统总生物多样性之间的假设关系。注意,jr轴是非定量的。这四条曲线说明了四种不同的情景,代表了农业管理对总生物多样性的不同影响,以及对保护的不同影响,特别是对生物多样性保护本身的影响。这个案例表明,最初的管理阶段对生物多样性的影响很小,在达到管理强度的某个相当关键的阶段之前,进一步的损失是逐渐的。在图11.2中,这被任意描绘为位于低强度和中等强度之间,但生物多样性迅速下降的关键阶段可能位于强度轴上的任何其他点。如果这种关系成立,那么生物多样性保护的规划活动应侧重于将管理强度维持在临界点以下,而不是以零管理战略为目标。这可以被认为是中间干扰假设的一个有趣的应用(Connell 1978)。Janzen(1973)提出,为了支持这一点,随意管理的农业生态系统实际上可能比未经管理的农业生态系统促进更多的物种多样性。例如,显示出以前维持生计的农业活动伤痕的森林似乎比那些缺乏这种干预的森林具有更高的物种多样性。
重要的是要记住,农业变化的模式是多样的。并不是像图11.2所暗示的那样,集约化必然来自于不受管理的森林,通过轮作,到固定间作,到集约化谷物种植。此外,也没有理由沿着强化轴的一个方向移动,强化就不能逆转。农业生态系统在特定的集约水平上确实可能向相反的方向转变。例如,哥斯达黎加以前的香蕉种植园最近已改为小型家庭维持生计的生产。因此,图11.2中并没有特别的变化路径,而是简单地观察到,管理强度的定性尺度可以大致有序,而生物多样性则是对这种有序的功能响应。
尽管如此,尽管图11.2中任何假设的曲线缺乏明确的证据,但还是有可能检测到一些变化模式在具体的例子中。
11.2.4变化模式
哥斯达黎加咖啡农业生态系统的转变动态为管理强度对生物多样性的影响提供了一个有益的例子。与其他主要的生态系统转型一样热带在美国,咖啡生产的转变涉及壮观的景观变化。这种转变的两个极端是传统系统和现代集约化咖啡单一栽培。前者遵循传统农林业的常见模式,有各种遮荫树种,经常点缀着果树,有时在咖啡灌木上方的森林“树冠”中相对密集地种植香蕉(Musa sp.)。咖啡本身往往是在单个咖啡树的水平上进行管理的,比如修剪会产生小的光隙,在其中种植木薯(Manihot esculenta)、山药(Dioscorea alata)或其他一年生作物。当一整群咖啡灌木被“翻新”(移除并重新种植新灌木)时,会产生一个更大的“光隙”,可能会种植玉米、大豆或其他需要光照的作物。因此,传统咖啡农场具有许多通常与森林相关的结构属性。
新的单一文化!该系统正在整个中美洲推广(Reynolds 1991;Babbar 1993)是完全不同的。所有遮荫树都被淘汰,传统的咖啡品种被新的耐太阳和较矮的品种所取代,这些品种在基因上是同质的。咖啡是按行或按块修剪的,严重依赖于农用化学品,特别是除草剂和化肥(icaf - mag 1989)。
这两个系统代表了咖啡管理系统连续体的两个极端,具有不同程度的复杂性。与集约化相关的植被变化在景观层面上是明显的,但在生物多样性方面更为微妙的变化则更为壮观。Nestel和Dickschen(1990)报告了传统系统中节肢动物的高度多样性,与无阴影的现代系统相比,Perfecto和他的同事(Perfecto和Snelling 1994;Perfecto和Vandermeer 1994)报告说,随着转变的进行,地面觅食蚂蚁的物种多样性显著减少。使用冠层杀虫雾法采集的树栖昆虫动物群初步样本表明,在该农业生态系统中可以发现昆虫的高度多样性(表11.1)。咖啡种植园遮荫树中鞘翅目和蚂蚁(膜翅目:蚁科)的多样性与其他人报道的相同数量级(Erwin and Scott 1980;Adis等人1984;Wilson 1987, 1988)中的甲虫和蚂蚁热带雨林的树木(表11.1)。请注意,采样的传统咖啡农场距离任何类似热带雨林的地方都有数百公里,实际上是现代无阴影咖啡海洋中的岛屿。虽然传统咖啡农业生态系统的甲虫和蚂蚁多样性与天然雨林的数字惊人地相似,但一旦系统现代化,它们就会迅速下降,这表明它们接近II型或IV型曲线。
自然系统向集约化单一栽培的转变导致土壤无脊椎动物和微生物多样性的丧失。随着能源、水和农用化学品投入的增加以维持生产,在自然系统中调节土壤生物过程的官能团被土壤肥力的机械和化学控制所取代。
图11.2所示,决定不同农业生态系统总生物多样性的关键因素是小气候、生境结构和食物资源。raybet雷竞技最新因此,人工林和家庭花园保留了许多天然森林的群落特征,而牧场群落在功能上与稀树草原相似。沿农业发展梯度的土壤生物群物种损失模式与图11.2中的曲线IV近似。然而,根据耕作制度和农业实践的类型,不同规模和不同功能的群体,从多样性较多到多样性较少的社区的拐点有所不同。在直接将森林转为密集种植短轮作作物的极端情况下,地表活动的大型无脊椎动物(千足虫、蚯蚓例如,甲虫),它们将落叶作为栖息地和食物资源(Lavelle et al. 1994)。随着土壤和凋落物有机质池的质量进一步减少,群落结构逐渐转变为占据缓冲土壤环境中的土壤孔隙的小生物(微节肢动物、线虫、原生动物、细菌、真菌),以及较大的生物,如蚯蚓和真菌白蚁由于大型生境的破坏,土壤中动物(螨类、岩尾类)的多样性随着耕作强度的增加而下降。
分类单元 |
树种 |
栖息地 |
国家 |
不。的物种 |
源 |
鞘翅目 |
Luehea seemannii |
湿润季节森林 |
巴拿马 |
继续阅读:红树林的生物多样性
这篇文章有用吗?

