红树林的生物多样性
生物多样性通常在三个层面上定义,即物种、种群和生态系统(Ray and McCormick 1992)。术语"生物多样性的组成部分已被推荐作为这几个层次的生态学参考,这个想法特别适合描述热带河口生态系统(Yanez-Arancibia et al. 1994)。热带河口的生物多样性组成部分可以指物种、生命史、栖息地和食物网中的联系的高度多样性,或者是陆海交界面陆地和海洋生态系统之间能量流动和营养循环的不同途径。此外,海岸地貌地貌和地球物理过程是有效调节河口生态系统特性的不同组成部分。河口环境条件的波动导致生物对栖息地利用的时空格局不同。红树林生态系统尤其如此,因为它们是与多种功能景观相互作用的开放系统,例如与陆源淡水、沿海海水、大气和沉积物-水界面的边界。Macnac(1968)使用术语“mangal”来指定沿海景观中的红树林生态系统的属性,而“mangrove-”则仅限于潮间带的特色种子植物。这一独特的术语描述红树林生态系统的特性,而很少用于红树林生态系统,强调有必要综合热带沿海生态系统的各种物理、化学和生物特征。
沿海地区多样的地貌可被视为红树林生态系统的生物多样性组成部分(图13.1)。这些区域可以划分为不同的地貌单元,以描述气候的影响
-
")
- t. Rwer-donúnaWd altoctuhonous它。以潮汐为主的异chihonus barcKi-hgoon (autoduhotwus)
")
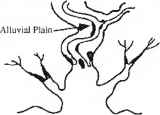
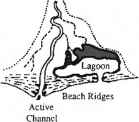
四、组合:河流、波浪为主。五、淹没基岩山谷
图13.1影响潮间带红树分布的五类基本地貌环境。重绘自《荆棘》(1982)
-

- 2-河流森林

-

- 4.-过度冲洗森林

5.——矮森林
图13.2红树林的生态类型(Lugo and Snedaker 1974)以及潮内带内这些生态类型的空间分布(来自Gilmore and Snedaker 1993,经John Wiley and Sons, Inc.许可转载)。箭头和罗马数字被用来根据Gilmore和Snedaker 1993年描述的红树林主要生境协会来定义七个特定空间行会的位置和运动。2树木的树冠;3底栖动物群落:潮溪沟群落;五、盆地群落;六、高地树木群落;7陆地群落地球物理过程对生态的影响红树的特征(刺1982)。红树林分布在五个基本的海岸环境中,这取决于地球物理能量的组合,包括降雨、河流流量、潮汐振幅、浑浊度和波浪能的相对影响(图13.1)。这五种环境环境都受到陆源物质输入的影响,而红树林也出现在碳酸盐台地,那里的环境环境以钙质为主沉积过程以及营养不良的条件(Woodroffe 1992)。这些碳酸盐台地群落的结构和功能与那些受陆源物质影响较大的红树林形成了有趣的对比。
一个地区的微观地形因素决定了土壤的许多水文和化学条件,这些条件控制着森林的地貌和分区模式。此外,潮间带的潮汐淹没频率可以影响繁殖体和物种的分布(Rabinowitz 1978年),尽管这种机制(“潮汐分选”)对森林结构的影响最近受到了质疑(Smith 1992年)。Lugo和Snedaker(1974)利用佛罗里达南部和加勒比地区红树林结构的局部格局,将红树林分为河流、边缘、盆地、吊床和矮林(图13.2)。红树林的这种生态分类还受到生物因素的影响,如对繁殖体的捕食(例如螃蟹)、幼苗对资源的不同利用以及树木的生理耐受性,这些因素决定了红树林的地貌和分区模式(Davis 1940;球1980;卢戈1980;Snedaker 1982;史密斯1992年)。这两种类型的分类系统,即地貌(图13.1)和生态(图13.2),代表了海岸景观的不同组织层次。它们一起可用来综合控制森林结构属性的不同尺度的环境因素(图13.3)。
树木的物种丰富度是红树林生态系统的另一个生物多样性组成部分(图13.4),上述环境设置和生物因素不仅影响不同地貌和生态类型红树林的形成,而且还可能控制物种丰富度(Smith 1992)。很明显,在一个大陆区域内,降雨量、温度和潮汐范围的变化可能对气候变化很重要红树林的多样性树(史密斯和杜克1987)。然而,有一些生物地理因素导致了物种丰富度的全球分布不平衡(Tomlinson 1986)。西半球(11种)的红树林树种多样性低于东半球(30多种)(图13.4)。这也导致沿旧大陆潮间带的分区模式比新热带地区的更简单的模式要复杂得多(例如Watson, 1928,与Davis, 1940;
-
 and ecological (local) factors that control the concentration of nutrient resources and stressors in soil")
- 图13.3基于全球、地貌(区域)和生态(局部)因素控制土壤养分资源和压力源浓度的分级分类系统,描述红树林结构和功能的不同模式
 and ecological (local) factors that control the concentration of nutrient resources and stressors in soil")
见查普曼1976)。目前,一般的概念模型已经改进,以解释特定大陆区域内的分区和森林结构的发展(Smith 1992;Gilmorc和Snedaker 1993),但由于缺乏对沿海森林发展的多种途径的了解,开发特定的生态模型来预测红树林物种丰富度和生态类型对土地使用或全球气候变化的响应仍然受到限制(Twilley 1995)。raybet雷竞技最新
红树林生态系统支持各种海洋和河口食物网,涉及数量异常庞大的动物物种(Macnae 1968;奥德姆和希尔德1972年;Yanez-Araneibia等人,1988;罗伯逊和
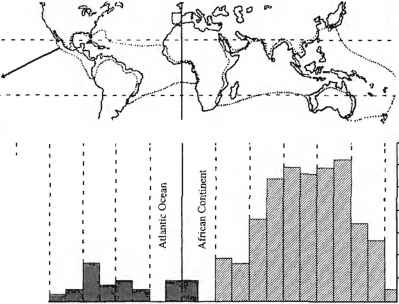
图13.4红树林的广义全球分布,包括所有物种的近似界限(上面板)和显示每经度15e红树林物种的近似数量的直方图(下面板)(来自Tomlinson 1986年)。经剑桥大学出版社许可转载)
120 90 60 30 WOE 30 60 90 120 150经度
西部红树林东部红树林
图13.4红树林的广义全球分布,包括所有物种的近似界限(上面板)和显示每经度15e红树林物种的近似数量的直方图(下面板)(来自Tomlinson 1986年)。经剑桥大学出版社许可转载)
120 90 60 30 WOE 30 60 90 120 150经度
西部红树林东部红树林
杜克大学1990)。输出的颗粒有机物(POM)支持了源自颗粒喂食者的食物网,而有时更大的可溶性(溶解)有机物(DOM)的输出形成了近岸异养微生物食物网的基础(Odum 1971;Alongi 1988;Snedaker 1989;Robertson et al. 1992)。许多利用红树林栖息地及其有机资源的鳍鱼和无脊椎动物物种也是近海地区的组成部分,这一现象表明了昼夜迁徙和季节性迁徙的复杂模式(cf. Thayer et al. 1987;Yanez-Arancibia等人,1988;Sasekumar et al. 1992)。除了海洋河口食物网和相关物种外,还有数量相对较多、种类较多的动物,从陆地昆虫到鸟类,它们生活在红树林植被中或直接以红树林植被为食。这些生物包括固着生物,如牡蛎和被衣动物,树食动物,如叶食动物和果食动物。 and ground-level seed predators. In consideration of the entire resident and casual faunal population in south Florida mangroves, Gilmore and Snedaker (1993) were able to recognize four distinct spatial guilds that may have well over an estimated 200 species, many of which are as yet uncataloged. In addition, Simberloff and Wilson (1969) documented over 200 species of insects in mangroves in the Florida Keys. For reference, the Florida mangroves consist of only three major tree species and one minor species of vascular plants. Based on these considerations, one can conclude that the low species richness of mangroves in Florida supports a disproportionately rich diversity of animals, the dimensions of which arc only now being documented. This same conclusion can be applied to other parts of the Caribbean (Ruetzler and Feller 1988; Bacon 1990; Feller 1993). Even though there is a global difference in species richness of mangrove trees between the east and west hemispheres, there does not seem to be a corresponding contrast in the functional diversity of the associated fauna. One exception is that Robertson and Blaber (1992) suggested that species richness of fish communities in the tropical Atlantic Ocean region was less than in the Indo-Pacific areas.
13.3森林结构与生态系统功能13.3.1红树林对养分动态的影响
红树林树冠层产生的垃圾影响循环无机营养物质在森林地面,以及有机物向邻近沿海水域的向外扩散(图13.5)(Odum and Heald 1972;特威利在。1986)。因此,红树林凋落物的动态,包括生产力、分解和输出,影响着红树林生态系统的养分和有机质收支(Twiliey 1988)。红树林是森林生态系统,许多陆地森林所描述的营养循环的生态功能也可能发生在这些潮内森林中。凋落物的数量和凋落物的质量(由C:N比值和木质素和多酚的浓度表示)有助于森林生态系统的营养动态(Aber和Mclillo 1982;麦克里洛在。1982)。因此,氮循环在森林的树冠与森林土壤中的营养动态相耦合,而这些受树木的特定物种营养生态的影响。测试这些反馈机制的研究将有助于了解树木生物多样性对红树林生态系统凋落物和养分动态的生态学意义。
在红树林生态系统的森林地面上,落叶层的积累可能是分解过程中养分固定的重要地点(图13.5;参见第13.3.2节)(Twiliey et at。1986)。落叶层中氮的浓度通常在森林地面的分解过程中增加(Heald 1969;Rice和Tenore 1981;特威利在。1986;日晚。
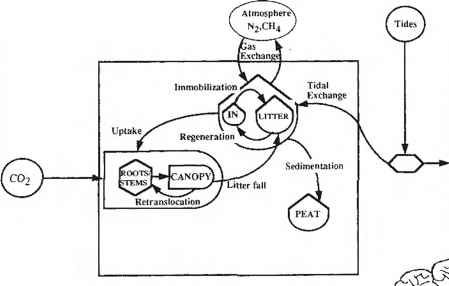
大气交换= AE固定= IM凋落物掉落= LF rcs = RT再生= RG沉积= SD潮汐交换= TE摄取= UT
继续阅读:红树林动物群与生态系统功能
这篇文章有用吗?
读者的问题
-
乔安3个月前
- 回复
-
哈桑一年前
- 回复
-
阿伊努人Tuikka一年前
- 回复
-
凯莉一年前
- 回复
-
休•贝尔一年前
- 回复
-
medhanie一年前
- 回复
-
紫水晶一年前
- 回复
-
珍妮特一年前
- 回复
-
任菲一年前
- 回复
-
阿齐兹达拉克一年前
- 回复
-
maddalena一年前
- 回复
-
纳塔莉亚一年前
- 回复
-
月见草一年前
- 回复
-
洋槐一年前
- 回复
-
曼科尔一年前
- 回复
-
Fiorenza一年前
- 回复
-
布兰科猪圈一年前
- 回复
-
kiros一年前
- 回复
-
丽娜一年前
- 回复
-
柯蒂斯一年前
- 回复