信息Ffm
7.4.3官能团
最近被广泛争论的一个话题是官能团(Walker 1992),其相关性可以从上面关于灌木营养循环的讨论中看到。功能类群的概念基于这样一个概念,即在分类上不同的生物体在功能上可能相似。一个明显连贯的功能群是与之形成共生关系的一组植物固氮细菌-这些最显著的是豆科植物(Rundel 1989),但它包括许多其他分类群。在太。固氮细菌主要有加利福尼亚的Ceanothus、Lotus和Cercocarpus属(Hanes 1977),南非的Apalathus和Psoralea属(Lamont 1983),智利的Trevoa trinervis属(Rundel 1983)以及西澳大利亚的许多种Acacia和Casurina属(Lamont 1983)。Blondel和Aronson(995)报告说,固氮剂在地中海盆地并不常见,但在法国南部和西班牙常绿橡树下形成密集林分的C’oriaria myrtifolia和杨梅属植物可能是重要的固氮剂(Rundel 1983)。豆科植物通常形成根结节与根瘤菌有关,而其他类群,如Casurina和Myrica会在受到放线菌感染时结瘤,而澳大利亚苏铁Macrozamia与蓝绿藻有关,形成环状固氮根(Lamont 1983)。
在澳大利亚西南部的eucaJpyt林地中,一组6到10种金合欢植物组成了可以被解释为氮固定物的功能群。由于它们的形态相似,而且都是固定氮,因此在组内具有明显的功能对等。然而,当考虑到它们对干扰的反应时,这种相似性就消失了。对这些物种子集的研究表明,不同物种的种子对高温有明显不同的反应,类似于火灾期间的预期反应(图7.4)。众所周知,某些物种的种子需要高温处理才能刺激发芽(Bell et at。1993),而其他物种的种子不耐高温。高温对金合欢植物有抑制作用,也有刺激作用,温度响应存在细微差异。因此,根据火灾的严重程度,不同的物种将被刺激发芽。因此,固氮群体内部的多样性为该群体在扰动强度变化时完全丧失提供了保险。在这个例子中,我们已经能够显示系统的组成(桉树和金合欢属),其结构(a木本植物有积累燃料周期的群落)和一些生态系统过程(火和营养物质的流动,以及遗传信息从一代到下一代的流动)。系统的机械部分的突发性属性,体现在一个其他紧密构成的功能组中,是整个系统能够随着时间的推移对不可预测性的火灾干扰模式作出反应的恢复力。可能还有其他同样重要的尚未被定量描述的涌现特性,如稳定性/自稳性、弹性、可塑性和可预测性。
显然,功能分组必须被视为相对的(Davis et al. 1994)。显然干旱-适应固氮剂与适应中等条件的固氮剂的用水组不同,且两者只能在有限的条件下相互替代(Hobbs et al. 1995b)。Jaksic等人(1993)和Fuentes等人(1995)所描述的在干旱的智利MTE中观察到的捕食者-猎物关系的动态很好地证明了功能组的相关性。在这项研究中,一组10种捕食者(4只鹰形鹰、4只猫头鹰和2只狐狸)与它们的猎物一起被监测,其中包括8种小型哺乳动物(7种啮齿动物和1种有袋动物)。在这些组中,动物被认为属于少数营养行业之一;被捕食的物种要么是谷类动物,昆虫动物,树叶动物或杂食动物(可能的偏好),而捕食者被分类为杂食动物或专属食肉动物。在研究的5年期间,年降雨量从58毫米到513毫米不等(长期平均为206毫米)。与这些干湿年份相关的是初级生产的低谷和高峰以及小哺乳动物种群规模。与预期相反的是,当高降水促进植物生长时,在假定的营养行会内的小型哺乳动物种群并没有同步爆发,而是只有羽叶虫(一种谷类动物)、橄榄木(一种谷类/杂食动物)和线虫(一种有袋类食虫动物)的种群爆发,而长尾Oryzomys longicaudaius(一种谷类动物)和长尾Akodon longipilis(一种食虫/杂食动物)的种群没有爆发。此外,在相对干旱的年份里,8种小型哺乳动物中有5种从研究地点消失。它们分别是长尾蛇(O. longicaudatus)、长尾蛇(A. longipilis)和三种叶食动物贝氏大柳(Abrocoma bennettii)、八爪蛇(Octodon dregus)和灰栗鼠(Chinchilla laigera)。 Fuentes et al. (1995) interpret these observations as evidence for little redundancy in the supposed guilds, and claim that these groups could not be functionally equivalent. The results therefore suggest that functional groups, should they exist for the prey species, cannot be defined in trophic terms alone.
另一方面,在捕食者的营养行业中,功能对等的迹象要强烈得多。两种食肉猫头鹰,Bubo virginianus和Tyto alba,以几乎相同的频率捕食同一物种,而杂食性协会包括两种假狐狸(P. culpaeus和P. gr ' sens),猫头鹰Athene cuunicularia和Glauci-dium nanum,以及猎鹰Falco sparverius,也表现出一致的相似的进食习惯。三种大型猎鹰(Buteo polyosoma, Geranoaetus melanoleucus和Parabuteo unieinctus)也一直是食肉动物。在贫瘠的年份里,当小型哺乳动物的数量下降到100只以下时,捕食物种开始消失,哈~虽然三只较大的猎鹰和三只猫头鹰的食肉饮食之间有合理的匹配,但前者首先从研究地点消失。就连杂食性猎鹰F. sparverius也比它的同类猫头鹰A. cuunicularia消失得更早。在猎物数量较低时留在研究地点的四种捕食者中,有三种(两只狐狸和猫头鹰G. nanum)是杂食动物,只有一种(猫头鹰T. alba)是食肉动物。在贫瘠的年份里,有6个物种离开了这个地方,其中5个是食肉动物。Fuentes等人(1995)认为,这种模式表明,在公认的公会之间存在高度的功能对等,但也承认,该系统尚未对公会内部的密度补偿进行“酸性测试”。
Wiens(1993)在对这一研究的综述中,对功能对等的相对性质作了进一步的解释。他解释说,在贫瘠的年份,鹰形物种的早期消失是一种功能组合的微调,以及在丰年,猫头鹰和猎鹰物种占据的广阔营养生态位被分离成独立和狭窄的生态位(图7.5)。就其逻辑结论而言,物种之间沿不同轴的功能属性分离最终归结为关于生态位的争论
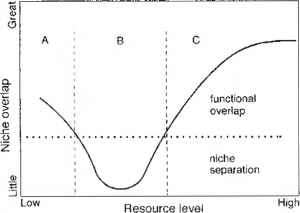
图7.5资源可用性与生态位空间重叠之间的关系,基于对智利捕食者猎物系统的观察,并证明了生态位空间重叠的相对性质生态位宽度.资源水平。A、B和C分别反映了被捕食物种的饥饿、专业化和机会主义条件。重绘自Wiens (1993)
图7.5资源可用性和生态位空间重叠之间的关系,基于对智利捕食者猎物系统的观察,并展示了生态位宽度的相对性质。资源水平。A、B和C分别反映了被捕食物种的饥饿、专业化和机会主义条件。重绘自Wiens (1993)
分化(sense Hutchinson 1958)。科迪(1986)已经优雅地证明了加州mte中这种生态位的存在。沿着一组横断面,这组横断面与沿海山脉的主轴相交(代表陡峭的环境梯度),另一组横断面与海岸平行(代表纬度环境变化的浅梯度),他确定了Ceanothus和Arctostaphylos不同属的物种更替。沿着这两组梯度,物种的替换序列相对陡峭的梯度较陡,较浅的梯度较浅,Cody(1986)解释为这两个被调查物种属的生态位分离的证据。
7.4.4楔石
绰号keystone可以应用于一个组件,有时也可以应用于一个系统的进程,删除它会对系统造成不成比例的变化(Lamont 1992;债券1993)。因此,基石质量是一个以特定方式将多样性与系统功能联系起来的概念,这表明物种丰富度的维持本身并不总是系统稳定性的可靠衡量标准,其他物种,甚至物种组合,可能依赖于假定的基石的存在。在澳大利亚对MTEs生物多样性的综述(Hobbs 1992)中,Lamont(1992)提出了对关键因素的多层次解释,利用自然系统中的相互作用来说明他的模型。在这个模型中,在一幅描绘为中世纪风格建筑的草图中,他指出了贾拉森林中三个层次的关键石:一级关键石只有一个物种依赖于它们;二级键石支持整个物种组合(例如,支持所有结瘤豆科植物的N2-fixmg细菌);三级基石是指没有它整个系统就会崩溃的基石。Lamont(1992)引用了一种主要的固氮植物——叶腹草(Gastrolobium bilobum)的例子,这种植物还能稳定土壤,为小型脊椎动物(如小型有袋动物Bettongia penicillata)提供食物和住所。G. bilobums种子的萌发依赖于一组外生菌根的作用,这些外生菌根反过来依赖于Bettongia的扩散,以及促进孢子萌发通过动物的肠道。据推测,如果没有G. bibobum, Jarrah森林系统将会崩溃。
最近关于蚂蚁传播种子的工作(Bond and Slingsby 1984;债券和股票1989;Bond el al. 1992)。超过1300种fynbos植物(20%的植物区系)产生的种子含有丰富的蛋白质,吸引了当地的蚂蚁。当地的蚂蚁习惯性地寻找种子,这些种子落在母体植物附近,然后把它们拖到它们的巢穴,在那里它们会被吃掉。然后,种子被丢弃在巢中,为它们提供保护,以抵御食谷动物和大火的热量。蚂蚁的作用是散播和保护种子,因此,蚂蚁的授粉可以被认为是许多fynbos植物继续生存所必需的关键过程。最近,外来的阿根廷蚂蚁Jridomyrmex humilis入侵了fynbos的部分地区,显示了这一关键过程的脆弱性。这种蚂蚁比本土的种子采集蚂蚁更小,但更具攻击性,并经常取代后者。阿根廷蚂蚁吃土壤表面的种子,不埋种子。 In fynbos invaded by this species, seedling regeneration of ant-dispersed plants after fire is much less successful than in uninvaded fynbos (Bond and Slingsby 1984). Besides threatening many fynbos plants with extinction, the collapse of this ant plant mutualism could have ecosystem-level effects since ant-dispersed plants are often dominant components of fynbos shrublands. Proteaccous species are generally deeper rooted than other members of fynbos communities (Higgins et al. 1987), and local extinction would probably therefore also induce a marked change in the hydrology of the host systems. Thus I. humilis may be considered a keystone invader with negative system impact.
7.4.5生物多样性及其对人类效用的支持
有许多例子说明生物多样性在生态系统的运作中可以发挥的作用。在某些情况下,组件本身的多样性或它们所提供的结构,对于自然或人为扰动影响所需的稳定性和恢复力可能是绝对必要的。然而,在某些情况下,生态系统多样性的改变可能会增强其对人类的效用。例如,在智利,蜂蜜产业的生产环节包括蜜蜂。蜜蜂是一种生命植物,从中获得蜂蜜生产的原料。这个系统已经被Varela el在。(1991)并在Fuentes el al.(1995)中报道,并对生物多样性可以发挥的作用提供了相当大的见解。首先,在一项调查中,蜜蜂全年收集的花粉表明,无论开花植物的种类有多少,蜜蜂只使用了其中的15种,这些植物在任何时候都多达90种。这表明按照第7.4.2节中提到的Vitousek和Hooper(1993)模型,多样性的功能饱和。第二个教训来自于蜜蜂收集的大部分花粉是由有限数量的植物物种贡献的事实(Varela et al. 199 i)。Galega officinalis (Fabaceae), Lithraea caustica (Anacardiaceae)和三个芸苔科成员(Hirschfeldia incana, Raphanus sativus和Rapistrum rugosum)属于这一类,在早期观察到花粉供应的季节性替换系列南半球夏季, 10月份大部分花粉由芸苔科植物提供
11 / 12月为L. caustica, 12 / 1月为G. officinalis (Fuentes et ai。1995)。在重要的花粉贡献者中,50%以上是引种植物(包括上面提到的芸苔科和野甘菊)。这表明,改变的多样性在智利中部生产供人类消费的蜂蜜的生态系统的功能中起着重要作用——还要记住,主要主角Apis mellifera也是一个引进物种。
南非fynbos地区也有类似的情况,在这种情况下,开普蜜蜂(A. mellifera capensis)是欧洲蜜蜂的fynbos种族(Hepburn and Jacot-Guiilarmod 1991),是本土蜜蜂,而许多为其提供花粉和花蜜的植物物种都是外来物种。开普蜜蜂使用的引进植物种类包括桉树、金合欢、柑橘类水果和落叶水果,以及许多草本物种(Anderson et al. 1983),导致了比没有它们的情况下更多产的蜂蜜产业。
7.4.6地中海盆地种内变异与系统功能
在地中海盆地,分类学上众所周知的物种群已被证明具有大量的种内变异。然而,将这种变异与功能联系起来一直很困难。例如,常见的牧草Dactylis glomerata由一个复合体组成,其中包括多达J 5个二倍体类型,三个四倍体和一个六倍体,后者仅限于北非(Lumaret 1988)。Stebbins和Zohary(1959)解释说,D. glomerata的四倍体形式的分化是温带和地中海类群的二倍体同源多倍体的结果,以及对不同类群的生态适应气候地区.地中海盆地的高环境异质性被认为是选择性的背后的力量该物种的MTE生态变异高于其范围内地形较为均匀的部分(Lumaret 1988)。对地中海条件的适应包括支持节水机制的形态特征,以及在整个夏季干旱期间保持种子。水关系的变化和本种的其他生理特征沿梯度水的压力或多或少地与四种不同酶系统的趋势相关(Roy和Lamaret 1987)。因此,种内变异可以被视为原料!对于进化来说,进化反过来提供了系统层面的功能可塑性,并确保了生态系统的持久性。
许多地中海植物的另一个重要特征是它们的组织中存在挥发性精油。众所周知的例子有百里香、薄荷、罗勒、欧芹、茴香、鼠尾草、迷迭香、薰衣草、香菜、牛至、芸香、月桂叶、艾草、胡芦巴、芝麻、藏红花、甘草、洋葱、青葱、香葱和大蒜。这些植物大多被分组在Lamiaceae, Apiaceae和Asteraceae。虽然这些物种显然对人类具有一定的功能价值,但含有挥发性芳香族化合物的植物在生态系统中的作用是复杂的,而且尚未完全了解。一方面,人们认为这些高度易燃的油与地中海盆地系统的火灾生态有关,但它们也可能:(i)是对食草动物、细菌和真菌的防御;(ii)通过抑制竞争对手的建立相互影响;(iii)模拟昆虫信息素吸引传粉者;(iv)通过提供抗蒸腾作用来减轻水压力(Margaris和Vokou 1982年)。胸腺属是研究得最好的芳香属之一,在种与种之间的含油量有显著差异,种与种之间也有遗传控制的差异。Gouyon等(1986)发现,寻常胸腺(Thymus vulgaris)中油分含量(化学型)的种内变异分布可能是由环境严格决定的。然而,他们也发现,在很短的距离内,化学多态有很高的周转,这构成了一个在时间上持续存在的马赛克。尽管人们对这一点了解甚少,但种内水平上的多样性显然与系统功能的维持有关。
7.4.7更新世食草动物在加利福尼亚古景观
在加利福尼亚,更新世末期,大型哺乳食草动物群发生了大规模的灭绝事件;在短短几百年的时间里,超过70%的属消失了。这显然是由于人类的影响导致了大型食草动物种群的减少(Martin 1984)。这导致了一连串的灭绝,包括营养链上的其他哺乳动物(Owen-Smith 1989)。认为大型食草动物的重要作用是维持景观多样性;这种动物性成分的丧失导致了其他食草动物所需栖息地的丧失,从而导致食肉动物食物资源的丧失,最终极大地改变了这些生态系统的功能(Keeley和Swift, 1995)。
7.4.8南非MTEs地层移位
直到最近,流行的观点是边界是固定的主要的植被南非开普南部和西南部的地层受到土壤因素和水分有效性的控制(见Cowling和Holmes 1992年的综述)。然而,最近的研究揭示了森林与fynbos、fynbos与草地、fynbos与雷诺斯特草原、雷诺斯特草原与草地之间边界的动态性质,以及自然植被与外来树木和灌木灌木丛之间的边界。在所有的情况下,改变干扰体制(特别是火灾和
-

- 图7.6 mte中连接系统过程与植被类型的反馈回路模型。该图指的是南非fynbos地区MTEs中观察到的自然植被动态和以外来入侵植物为主的动态,如表7.4所述

放牧强度)导致地层边界的变化(Richardson et al. 1995)。
Cape区系的主要自然植被和外来植被的生态系统功能,如净初级生产力和常住植物量、产水、火行为、火引起的土壤拒水性、产沙量和养分循环都有很大差异。这些差异可归因于组合的结构和功能特征,而不是这些地层本身的生物多样性(Richardson et al. 1995)。不断变化的土地利用模式能够并且确实影响CFR主要地层之间的边界。物种丰富的fynbos被物种贫乏的原生森林或少数外来树种的林分所取代,对生态系统功能有重大影响。这种植被变化通常是快速和不可逆的(Bond and Richardson 1990),其影响是普遍的(图7.6)。例如,最近来自邻近生物群落的许多鸟类入侵了该地区,这表明人类引起的植被变化对许多生态系统特征造成了深刻的改变。有意或自然发生的地层转移为研究生物多样性变化对生态系统功能的影响提供了极好的机会(表7.3)。
继续阅读:草原函数模型
这篇文章有用吗?


