全球土壤有机碳库
迈克尔的鸟
澳大利亚国立大学,
堪培拉,澳大利亚
Hana Santruckova
南波西米亚大学,
Ceske Budijovice,捷克共和国
约翰·劳埃德
马克斯·普朗克研究所
生物地球化学,lena,德国
埃尔Veenendaal
哈里奥本海默奥卡万戈
博茨瓦纳芒市研究中心
1.简介:土壤碳池与全球变化185
2.影响因素土壤分布有机碳186
3.SOC池的全局变化
4.现有SOC观测数据的局限性
5.分层抽样法
6.结论:《沙世界》和《粘土世界》197
引用197年
1.引言:土壤碳库与全球变化
对全球土壤有机碳(SOC)库规模的估计范围在700 Pg (Bolin, 1970)和2946 Pg (Bohn, 1976)之间,目前普遍认为约1500 Gt的值是最合适的(表1)。这个值被认为是陆地碳库总量的一半(例如,Townsend等人,1995)和三分之二(例如,Trumbore等人,1996)之间。
SOC库在调节全球碳循环的人为变化中发挥着重要作用。一方面,人类活动,如土地清理、农业和生物质燃烧的铅到来自有机碳池的大量二氧化碳排放,这种排放往往在对常住生物量的初始扰动停止后很长时间内仍会继续。相反,由于光合作用的增强,陆地生物圈被认为正在隔离-■1 -2 Pg/年的人为CO(“缺失的碳汇”)碳固定(例如,Dixon et al., 1994)。虽然常住生物量被认为是平衡全球人为CO预算所需的增强吸收的原因,但SOC库被认为为大部分这种碳提供了长期的瞬态汇(Smith和Shuggart, 1993)。这是由于有机碳库需要相对较长的时间来建立一个新的平衡,并从现有生物量向土壤输送碳的速率提高。
尽管有机碳库在全球碳循环中起着主要作用,但有机碳库内部以及与其他主要全球碳库之间的碳交换动态碳水库约束差(Townsend等人,1995;Tans等人,1990;Trumbore, 1993;Fung et al, 1997)。主要的不确定性仍然围绕着有机碳池的大小,有机碳池存储由活生物量隔离的额外碳的能力,以及有机碳池对气候变化的响应。raybet雷竞技最新要减少这些不确定性,就需要对土壤碳库的大小、土壤碳库的速率和通量进行更可靠的估计,并制定S13C和AI4C测量所给出的额外限制(Fung等人,1997年)。
上述陈述暗示,需要能够更好地预测土壤基质类型、地貌和气候等SOC库的空间变化,并根据一致的全球观测数据库评估此类预测的准确性。raybet雷竞技最新《京都议定书》具有约束力的条款以及未来国家间碳交易的可能性,进一步迫切需要建立可核查的碳储量清单和碳流入和流出SOC库(最大但最不了解的陆地碳库)的通量(IGBP陆地碳工作组,1998年)。雷竞技手机版app
本章探讨了有机碳库观测研究的现状,土壤碳库的全球趋势,以及观测研究与有机碳库碳交换动态建模工作之间的关系,并提出了如何建立模型的建议
研究 |
土壤碳(Pg) |
柏林(1970) |
700 |
博翰(1982) |
2946 |
Baes et nl。(1977) |
1080 |
巴齐列维奇(1974) |
1392 |
施莱辛格(1977) |
1456 |
艾特杰等。(1979) |
2070 |
Post等人(1982) |
1395 |
Kswaran等人(1993) |
1 576 |
Baljes (1996) |
1500 |
观测和模拟研究可能会更好地结合起来,以改善围绕SOC池动力学的主要不确定性。
2.影响土壤有机碳分布的因素
Jenny(1941)首先阐明了可能影响土壤有机碳含量的因素,尽管之前已经确定了土壤、植被和气候之间的广泛关系。raybet雷竞技最新表2还利用了后期研究的数据,列出了控制SOC库存的主要因素。
raybet雷竞技最新气候(温度和降水)通过控制从活生物量进入土壤的输入水平,对全球范围内的有机碳(SOC)产生重大影响土壤。raybet雷竞技最新气候也会影响
温度 |
|
降水、蒸发 |
|
土壤质地(母质/时间) |
|
Geomorphoiogy |
|
排水/高程 |
|
坡 |
|
微环境 |
|
营养状况(母质/时间) |
|
自然干扰 |
|
火 |
|
干旱 |
|
昆虫和疾病 |
|
风倒 |
|
放牧压力 |
|
人为的干扰 |
|
清除/植树造林 |
|
生物质燃烧 |
|
农业 |
|
放牧压力 |
|
raybet雷竞技最新气候变化 |
|
有限公司/ N施肥 |
输送到土壤中的碳通过有机碳池循环,最终通过微生物生物量呼吸回大气,或者作为溶解的有机碳从剖面上损失(图1)。气候与其他因素结合,控制着初始凋落物质量(氮含量、木质素含量等;raybet雷竞技最新Melillo等人,1982)以及在有机碳池中改变有机碳性质的过程。raybet雷竞技最新气候通过影响光照和有效生物扰动的效率和深度,通过土壤剖面影响有机碳分布(例如,Holt和Coventry, 1990),是一个关键方面
纬度的腰带
高度的腰带
纬度的腰带
高度的腰带
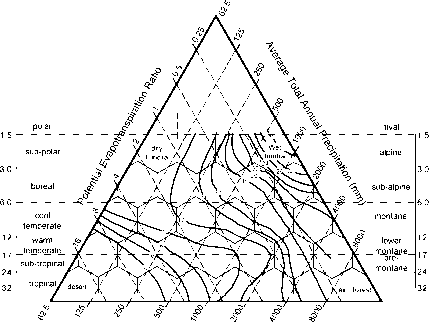
图1 Holdridge生命区中以kg/nr1为单位的有机碳分布(Post et ai, 1982)。
或影响生产速度和土壤基质矿物学(例如,Goh等,1976)。还应注意的是,时间在决定土壤基质性质方面起着重要作用,如成土过程在时间尺度上进行操作,从最近冲积沉积物上的成土作用的年份,到大陆深部风化作用的数百万年克拉通.
在局部尺度上(即对于特定的气候),其他几个因素调节着SOC在整个景观中的分布raybet雷竞技最新。在这个尺度上最重要的是土壤质地(Parton et al., 1987),这是一个与其他参数密切相关的变量,如基岩类型、养分状况(阳离子交换能力)、持水能力、光照和生物扰动率、根系穿透阻力以及支持好氧微生物呼吸的氧气有效性。方便的是,这些变量往往以这样一种方式耦合,土壤质地成为所有这些变量的有用代理,SOC水平通常随着土壤基质颗粒尺寸的减小而增加。
地貌通过决定沉积物和有机碳的侵蚀/增加速率以及水/营养物质的获取,以及通过提供当地微环境(例如,气候或火灾保护)来控制土壤碳水平,这些微环境允许植被类型的发展,以不同的速度隔离和循环碳。局部微环境也可以改变微生物呼吸速率。
近几十年来,由于土地利用变化在决定陆地碳源/汇与大气二氧化碳库之间的转移量方面发挥了重要作用,各种自然和人为干扰在修改有机碳库存方面的作用越来越受到关注。一些干扰,如砍伐森林/伐木、农业和放牧活动,显然是人为的,而其他干扰,如风力、极端气候(干旱等)、虫害和疾病,则更直接归因于自然原因。在诸如生物质燃烧之类的扰动情况下,通常更难将其归结为独特的人为或自然原因,而人为气候变化的可能性意味着所有上述扰动现在可能部分与人类活动有关。raybet雷竞技最新从这个意义上讲,C02/氮肥效应和造林也可归为影响有机碳池的人为扰动。
上述扰动对有机碳库的最直接影响是改变碳传递到土壤的速率,要么改变活生物量固碳的速率,要么将活生物量固碳直接或间接转移回大气。扰动还可以改变微生物呼吸作用使碳返回大气的速率。
在许多情况下,扰动会导致当地植被和土壤结构的长期变化,这意味着在扰动维持期间以及扰动停止后建立新的平衡期间,当地有机碳库既可以作为大气的来源,也可以作为大气的汇。因此,扰动可导致有机碳储量的永久变化和扰动区域碳通量的瞬时变化。因此,在考虑SOC池在扰动期间和扰动之后的响应时,时间是一个重要的变量。
所有上述变量之间可能存在的复杂相互作用是围绕土壤碳库内外碳交换动态存在许多不确定性的根本原因之一。
3.SOC池的全局变化
在来自不同气候的土壤之间的比较研究中,已经观察到有机碳库中可预测的变化(Jenny, 1961;raybet雷竞技最新Trumbore, 1993)或对比土壤质地类型(Parton等人,1987),沿气候梯度的横断面(Townsend等人,1995),跨土壤时间/地形序列的横断面(Goh等人,1976;),以及受干扰地区和未受干扰地区之间的比较(Townsend等人,1995;Desjardin et al, 1993)。此外,一些研究试图研究欧洲大陆的SOC趋势(例如,Spain等人,1983;Moraes等人,1995年)或全球(例如Post等人,1982年)规模,使用从文献来源编译的数据,已经能够推断SOC库存的主要趋势raybet雷竞技最新气候和土壤类型,但通常在数据中有很多分散。
基于对有机碳池行为的理解,已经开发了诸如Rothamsted (Jenkinson和Rayner, 1977)和Century (Parton等人,1993)等模型,可以将区域验证研究的结果外推到全球范围(Schimel等人,1994)。这些模型将有机碳池划分为3到5个池,循环时间从数年到数千年不等,给定土壤结构下这些池的大小由气候驱动的植物碳输入、养分、微生物呼吸和DOC浸出之间的相互作用决定(图2)。在某些情况下,这些模型已经过测试raybet雷竞技最新
-
.")
- 图2世纪模型中的碳流动(Parton et al., 1987)。
.")
表3土壤碳储量观测趋势 |
|
趋势 |
参考 |
raybet雷竞技最新Climate-specific趋势 |
|
SOC周转率随降低温度(T)及雨量(ppt) |
特朗博等人(1996) |
Trumbore (1993) |
|
表层土壤总有机碳比例随T(和ppt?)的降低而增加。 |
Zinke等人(1986) |
西班牙等等。(1983) |
|
微生物呼吸速率随T的降低而降低(和ppt?) |
劳埃德和泰勒(1994) |
瑞奇和施莱辛格(1992) |
|
SOC库存随着T的增加而减少(在恒定ppt下) |
Poster al. (1982) |
珍妮(1980) |
|
SOC存量随ppt的增加而增加(T不变) |
Post etal (1982) |
珍妮(1980) |
|
S] 'C在C的表面SOC中,仿生学随T的降低而增加 |
伯德等人(1996) |
草性生物群落表层有机碳<5I3C随T的降低而降低 |
Bird and Pousai (1997) |
L特定位置的趋势 |
|
粗纹理土壤的有机碳储量一般低于线纹理土壤 |
帕顿等人(1987) |
Schimel et al. (1994) |
|
SOC槽通常在地势较低的位置(山谷)较高 |
马洛等人(1974) |
西班牙等等。(1983) |
|
扰动通常会导致有机碳存量和通量的变化 |
达赖和迈耶(1986) |
Harrison等人(1995) |
|
SOC存量(每cm*)通常随着剖面深度的增加而减少 |
西班牙胎儿(1983) |
Desjardin等等。(1993) |
|
SOC的表观“年龄”通常随着剖面深度的增加而增加 |
汤森等等。(1995) |
Desjardin等等。(1993) |
|
在草木混合系统中,树下有机碳储量一般较高 |
Kellman (1979) |
Bird and Pousai (1997) |
|
木质组织的有机碳比非木质组织的有机碳有更长的周转时间 |
帕顿等人(1987) |
Bird and Pousai (1997) |
|
在排水良好的土壤中,SOC 8’*C值一般随深度增加而增加 |
格林等人(1996) |
Kruil和retaliation(印刷中) |
|
在排水不良的土壤中,SOC§I3C值一般随深度的增加而降低 |
Agren等等。(1996) |
Krull和retaliation(印刷中) |
|
在周期性排水不良的土壤中,有机碳值随深度而变化 |
Agren等等。(1996) |
Krull和retaliation(印刷中) |
|
细粒粘土有机碳比粗粒颗粒有机碳具有更高的SL1C |
Desjardin等等。(1993) |
Bird and Pousai (1997) |
|
细粒有机碳比粗粒有机碳在土壤中的停留时间长 |
布亚诺夫斯基等人(1994) |
在C,/C4混合体系中,SOC <5I3C随粒径的减小而增大 |
Bird and Pousai (1997) |
在所有站点,模拟SOC库存与观测SOC库存之间的一致性小于25% (Parton et ai, 1993)的全球分布现场站点的观测数据。
表3总结了前一节中所述因素相互作用导致的全球SOC趋势,并通过上述研究类型加以阐明。
土壤碳储量的主要全球趋势与气候和土壤质地之间的相互作用关系最为密切。raybet雷竞技最新因此,在潮湿、低pet(潜在évapotranspiration)的热带/亚热带地区,由于植物产量高,SOC库存往往较高,而在亚热带地区,由于微生物再矿化率相对较低。相反,炎热干燥地区的有机碳存量较低,因为植物产量低于微生物的再矿化效率(Post等,1982年)。在任何地区,由于有机碳和粘土颗粒之间的有机矿物相互作用,在这种气候控制的基础上,细纹理土壤往往比粗纹理土壤具有更高的碳储量化学稳定较高比例的SOC (Mayer, 1994;Skjemstad等人,1993;1996)。SOC、气候和土壤质地之间的广泛关系如图3所示(另见颜色插入)。raybet雷竞技最新
由于碳和营养物质的下坡运动以及更可靠的获取途径,局部地貌效应往往会导致地势较低的地方碳密度较高地下水和地表水.沿河道的碳储量往往较高,在火灾易发地区,在免于火灾的局部地形洼地,碳储量往往较高。有机碳地貌局部化的极端例子是泥炭沼泽的发育。
景观中树木的分布对碳储量有显著影响,树冠下的碳储量通常较高(例如,Liski, 1996)。树木分布的影响在稀树草原生态系统中尤为明显(Kellman, 1979;伯德等人,出版中;Bird and Pousai, 1997)。分布
0 14 28 42 58 70
潜在平均土壤碳周转率图3利用Century模型的温度和土壤质地关系推断出的全球尺度潜在平均土壤碳周转率(Schimel et al., 1994)。参见颜色插入。
热带稀树草原内树木的数量以及热带稀树草原/林地/森林边界的位置本身是气候、火灾频率、土壤类型、地貌和放牧压力之间复杂相互作用的结果(例如,Archer, 1990)。raybet雷竞技最新
在局部尺度上,通常可以在单一土壤剖面中确定有机碳的几种一般趋势。由于向土壤剖面输入的碳大部分来自于上覆的常绿生物量,土壤有机碳含量通常沿剖面下降,近似为对数-对数函数(Zinke et al., 1986)。高达50%的1米有机碳储量可能出现在剖面的顶部20厘米,这里的表面被视为矿物土壤层的顶部。碳在上层土壤中集中的程度是土壤类型、生根深度和气候的函数。raybet雷竞技最新密集、深度的生物扰动,如大多数热带地区的特征,与生物扰动最小的寒冷地区相比,导致碳储量随深度的逐渐下降,而永久冻土可能会限制SOC的向下移动。草地土壤的有机碳在较深的土层中所占的比例往往高于类似的林地,这可能与草地生态系统的生根较深有关。
随着温度和(或)降水的降低,土壤中碳的平均周转时间(以14C表示)通常会增加(Trumbore, 1993),这反映在微生物呼吸速率的减慢上(Raich和Schlesinger, 1992)。在单个土壤剖面中,土壤碳的14c年龄往往随着土壤剖面的深度而增加(Townsend等,1995;Desjardin et al., 1993)和颗粒尺寸减小(Buyanovsky et al., 1994),反映了细颗粒和深层土壤中旧难降解碳的相对比例增加。
许多研究研究了土壤中SMC分布的控制作用。对有机碳S”C组成的主要控制是从活生物量输送到有机碳池的碳的S13C值。陆地生态系统中C和C4植被的分布控制了很大的差异,而这又由气候决定(Lloyd和Farquhar, 1994)。raybet雷竞技最新较小的差异由海拔(Bird et al., 1994)、土壤水分有效性(Stewart et al., 1995)、辐照度(Ehrlinger et al., 1986)和封闭冠层中呼吸CO的再利用程度等因素决定(Van der Merwe and Medina, 1989)。
碳在被植被同化后可以在有机碳库中保留很长一段时间。这意味着,由于工业化以来化石燃料的燃烧导致大气中<5’3C值的下降,并不是SOC库中的所有碳都与现代大气的同位素组成处于平衡状态(Freidli et al, 1986;Bird et al, 1996)。因此,“旧”碳将比新碳富集1.5%。在计算从有机碳池中呼吸到大气中的二氧化碳的5’3C值时,需要考虑这种“陆地苏斯效应”(Fung et al, 1997)。
碳的代谢,一旦进入土壤微生物和动物进入有机碳池,就会逐渐改变有机碳的I3C值。在决定代谢过程中剩余碳的<5I3C值时,有两个主要过程相互竞争。选择性利用富含营养和能量的化合物,如糖和蛋白质,往往会增加剩余碳中木质素等成分的相对比例,这些化合物的<5I3C值低于其组成部分的总体生物量(Benner et al, 1987)。相反,伴随着代谢的动力学分馏效应倾向于有利于1JC的呼吸,从而增加了划分为微生物生物量的碳的¿>I3C值(Macko和Estep, 1984;Blair等人,1985年),并最终进入“缓慢”和“被动”SOC池。
由Agren等人(1996)提出的模型将有机碳中<5I3C的碳与作为微生物呼吸底物的凋落物的初始“质量”联系起来,低质量凋落物降解碳的<5I3C值逐渐变化为小或负,而高质量凋落物降解碳的<5I3C值逐渐变化为高且正。Krull和Retallack(已出版)提出,在土壤中观察到的伴随退化的分馏可以分为两大类。在通风不良的土壤中,当好氧呼吸被阻滞时,有机物会积累,对某些化合物的选择性利用会导致剩余碳的13C值降低,这是由于较难处理的低l3C木质素化合物的比例相对增加。在透气性好的土壤中,为动态的同位素分馏占主导地位,剩余的碳富集在13C中。
另一个尚未被考虑的机制是大型动物在有机物分解中的作用。例如,在季节性干燥的热带地区,据估计20%的有机物分解来自白蚁的作用(Holt和Coventry, 1990)。由于甲烷是白蚁巢穴中有机质分解的丰富产物,因此剩余的有机碳残留物可能在l3C中大量富集。
土壤中微生物代谢的部分“稳定”产物的浓度,加上自工业化以来大气二氧化碳SI3C值的下降,意味着旧的、难降解的、粘土相关的碳比粗颗粒碳具有更高的SI3C值(Kracht和Bird,在综述中)。在热带稀树草原,这种SI3C值随颗粒尺寸减小而增加的趋势,因c4衍生碳在细颗粒尺寸部分的优先积累而得到增强(Bird和Pousai, 1997)。
在区域/全球层面,关于SOC§13C值变化的观察性研究相对较少。Bird等人(1996)证实了森林土壤中表层有机碳的I3C值存在纬度梯度,热带森林土壤的I3C值比高纬度森林土壤的I3C值低大约l-1.5%。这归因于土壤中碳转换时间的温度梯度(“陆地苏斯效应”),也可能归因于~0.5%的温度效应(Bird和Pousai, 1997)。
Bird和Pousai(1997)发现,贯穿澳大利亚北部的降雨样带上的§13C值和碳含量可以用与季风森林一端、热带草原另一端以及分布在这两个端元之间的热带草原样本的对数线性关系来描述。
4.现有SOC观测数据的局限性
“由于采样深度和分析方法不一致,不可能对所有样本进行直接比较。”(Spain et al, 1983)
尽管在前面所述的SOC行为的控制因素方面取得了许多进展,但在全球范围内对SOC池行为进行精确预测的主要障碍是上述引用。几乎所有试图比较或汇编已发表研究结果的研究都出现了类似的说法。
这些问题在很大程度上源于许多早期的个体研究,这些研究的目的与当前全球变化研究的兴趣无关,因此,所采用的抽样策略对于构建全球SOC库存或SOC动态建模都不是最优的。此外,除了确认碳储量的总趋势外,现有的全球有机碳库存和上一节讨论的趋势并没有提供可用于验证有机碳动态模型的数据。
这些问题可以分为三种“类型”——定义、抽样和分析。关于土壤碳的定义存在一个重大问题。许多研究明确区分了“矿物土壤”、有机“o层”和“凋落物”,大多数对有机碳池大小的估计仅涉及矿物土壤中有机碳池的大小。然而,在冷温带地区,许多土壤的枯落物和o层中存在很大一部分有机碳。这种碳可能已经“死亡”了一个多世纪。从某种意义上说,这些物质中的大部分都可以被视为有机碳,因为它们是处于可变分解状态的死有机碳,为活的根状生物量提供了营养来源,并为微生物代谢提供了底物。
对于全球变化需求,SOC的一个更合适的定义可能是SOC表示从陆地表面向下(即传统凋落物层的顶部)且尺寸不大于2毫米的所有死碳,包括存在于凋落物和o层中的这些碳。因此,所有大于2毫米的材料都被认为是枯落物,这是土壤科学中常用的尺寸,是一种容易区分且易于量化的方法。目前凋落物和o层之间的区别是模糊的,因为实际上在凋落物、o层和矿物土壤中的碳之间存在着连续统一体。目前的区分也不能解释最近枯死的根衍生的地下凋落物的存在,其周转动态可能比矿物土壤更类似于地表凋落物。
另一个定义问题涉及许多研究中仅仅根据土壤类型来描述采样地点。
这可能再次用于土壤成因研究,并允许对可能控制SOC动态的因素进行一些推断。然而,它没有提供可量化的结构和地貌信息,而这些信息对于确定有机碳动力学至关重要,并作为建模研究的输入(Parton ct al., 1987)。许多研究也没有分别量化来自不同来源的碳输入。在森林中,当地的碳储量因树木的分布而有很大差异(Liski, 1996)。在混合热带稀树草原生态系统中,来自树源和草源的碳输入并不一定是相等的,并且会随着每种来源的比例而在空间上有所不同(这一问题在热带稀树草原SOC的§I3C值方面尤为突出)。
在比较不同研究的结果时,缺乏商定的抽样方案是一个主要问题。个别研究从不同深度增量采样到不同深度(或按层位),在许多情况下不包括土壤密度数据。在没有土壤密度数据的情况下,作者尝试使用各种推理技术从%C数据估计碳密度。虽然这可以提供碳密度的粗略估计,但Bird(1998)发现,放牧对类似玄武岩来源的土壤的影响之一是使0- 5厘米区间的土壤密度增加了25-35%,任何试图从单一的土壤密度近似估计中估算碳密度的尝试都会在计算碳密度时引入类似的误差。
土坑的一般使用意味着样品和容重测量通常在样品间隔的中点水平地进入坑壁,而不是通过收集整个深度间隔;如果碳在深度上的分布不均匀,这可能会带来(较小的)偏差。此外,关于样品是否筛分,是否/如何考虑土壤碳酸盐,文献中几乎没有一致性。
通过使用不同的分析技术,在先前发表的数据中引入了进一步的大不确定性。在高温下(以各种形式)在氧气中燃烧是目前确定土壤样品中碳丰度的首选方法。然而,许多区域和全球SOC库存研究所基于的大部分文献数据也包括使用Walkley-Black生成的数据湿法氧化或“点火损失”技术(例如,Moraes et al., 1995)。
人们早就知道,沃克-布莱克技术低估了有机碳总量,如果试图弥补这一事实,通常会将测量值乘以1.3倍。这个因子似乎来自沃克利和布莱克(1934)的原始工作,代表了七个土壤样品分析中碳的平均低估,其中为单个样品计算的因子范围在1.16到1.66之间。1.3的值已被广泛应用(例如,Little ct, 1962),但由于Walkley-Black氧化法不测量“顽固性”碳,可以预期,这种顽固性碳在土壤中的比例将高度可变,这取决于从立木生物量、火灾频率和其他土壤参数传递到土壤的碳的性质。
点火损失衡量的是“有机物”,一个单一的不变因子(一般为0.58,范布莱梅伦因子)通常乘以观察到的有机物含量,以计算样品中碳的百分比。然而,众所周知,例如,这一因子是可变的,在热带地区可低至0.45 (Burringh, 1984)。这个因素的不确定性将直接导致计算出的碳密度的不确定性。
上述讨论表明,根据用于估计同一地点碳储量的采样和分析方案,结果可能很容易相差10-20%。除了这种不确定性,还必须加上SOC池在所有空间尺度上固有的高变异性,以响应前面章节中概述的因素。
与活的植被不同,目前还没有办法远程监测SOC存量。SOC储量可以通过SOC模型与来自遥感数据的NPP估算相耦合来估算,但SOC观测数据的收集方式通常无法提供验证模型结果的手段。
就目前对有机碳储层碳同位素组成的了解而言,情况更加困难。这一领域的工作很少有例外(例如,Bird等人,1996;Bird和Pousai, 1997)专注于使用碳同位素作为局部地区碳动态和植被变化的示踪剂。目前迫切需要能够用于对有机碳库的碳同位素组成以及从有机碳库到大气的碳通量进行观测估计的技术。
现在有几种模型可以进行预测同位素分馏(例如,Lloyd和Farquhar, 1994),当结合大气同位素组成的测量时,这一知识对源/汇分布提供了重要的约束(Cias等人,1995;Fung等,1997;Bakwin et al, 1998)。
目前还没有办法用区域观测地球同位素数据更好地约束或检验这些模式结果。由于有机碳库整合了当地植被数年至数年的同位素特征,如果该特征能够充分地与退化的同位素效应(例如,Agren et al, 1996)和陆地“Seuss效应”(Bird et al, 1996;Fung et al, 1997)。
需要一种新的有机碳数据类别,以一致的方式在全球范围内收集,从而可以直接比较大范围气候条件下的结果,更好地与遥感植被指数相结合,并且更适合于全球碳循环动态模型和有机碳动态模型的验证。
5.分层抽样方法
前一节的讨论强调了试图利用以前发表的数据以足够的分辨率获得SOC池的全球观测趋势的固有问题,以便用于解决全球变化研究中的主要问题,例如“缺失汇”的位置。《京都议定书》的政治要求也可能要求对SOC池的规模和SOC池的通量进行比目前可能的更准确和可验证的估计。
Parton等人(1993)的方法是朝着这个方向迈出的有益一步,保持生物群落不变(使用全球分布的11个草地站点),并允许气候和土壤质地变化。raybet雷竞技最新然而,为了更好地定义全球趋势,一个更有指导意义的方法可能是保持土壤质地不变,并允许气候(从而允许植被)变化。raybet雷竞技最新因此,在任何特定气候下,砂质土壤上未受干扰的植被将提供一个“低”端元,而玄武岩衍生(或类似)土壤上的植被将为SOC库存和通量(包括同位素)提供一个“高”端元。raybet雷竞技最新然后,这两种估计可酌情混合,根据土壤质地分布的知识,提供给定地区的区域储存和通量的估计。
末次冰期末期的植被风沙体在世界上许多地方都很常见,古河道和古河道中的沙质沉积物也是如此冰川沉积物.同样,来自玄武岩(或侵入性等效物)的细质地土壤在全球范围内普遍分布。值得注意的是,沙质土壤比质地细腻的土壤具有显著的优势,因为它们通常可以自由排水,而且一旦土壤受到取样干扰,SOC的物理或化学保护的范围很小,这可能会使结果的解释复杂化。
这种方法忽略了二次地形效应,但是可以通过将采样限制在局部地形较高的位置来避免这些影响。因此,这种方法不能应对局部影响,例如泥炭的出现或水道周围不同的SOC动态,但目前任何其他方法都不能。
Holdridge(1947)提供了一个简单的气候分类在植被方面降雨和降水将世界划分为30个生活区。虽然这种分类已经被最近的工作所取代,但它可以作为覆盖全球可能需要的样本区域数量的指南,即120个地点(每个生命区域中重复的细纹理和粗纹理地点)。
在试图将最近的有机碳(0-5厘米)与深层土层(5-30厘米和30-100厘米)部分分离的每个地点,必须采用一致的采样深度。在温带地区,
HU-1
HU-1
HU-2
e 300
♦hu1 5- 30g□hu1 5- 30t ahu10 0- 5g xhu10 0- 5t
0.70 0.90 1.10 1.30 1.50干土密度[g cc'1]
1.70
HU-2
E 300
♦hu2 5- 30g□hu2 5- 30t a hu2 0- 5g xhu2 0- 5t
干土密度[g cc1]
1.70
图4土壤碳密度与土壤容重之间的关系,以及由这些数据平均计算出的库存量。这两个地点位于北昆士兰Hughenden-Charters Towers地区未开垦的玄武岩来源的土壤上(Bird, 1998),每个地点的样品都是沿着大约1000米的横断面收集的。采样深度分别为0-5cm和5-30cm。由于植被是树-草马赛克,总样本的一半是在距离树木半冠距离处(- T)收集的,另一半是在距离当地树木大约相等的距离处(- G)收集的。在现场清单的构建中,T和- G样本的总数根据现场估计的冠(- T)覆盖率加权。
如果存在较厚的表层有机层,则可能需要分别对其进行量化,并开始在矿物土壤的顶部进行深度采样。下面的所有讨论都将土壤表面定义为常规“凋落物”水平面的顶部。
如果树木被广泛分离,或者在生物群落中树木和草都存在,则必须从每个类别中收集一组单独的样本,因为树木分布对SOC池中的局部变异性有重大影响,如前几节所述。所有样品必须进行粒度分馏,以提供解释所观察到的变化和作为模型输入所需的纹理信息。
即使试图以上述方式控制变量,如果取样仅限于几个土坑,仍然会有局部的可变性,不能包括在内。Liski(1996)认为,对于北方森林在芬兰的砂质基底上,为获得的平均值的10%置信区间,至少需要30个样本碳排放清单,对于更加异质的树-草生态系统,所需的样本数量可能比这更高。Carter et al.(1998)发现,需要从昆士兰(澳大利亚)的1°X 1°网格单元中抽取15个随机土壤样本,才能将单元的平均有机碳含量定义为与“真实”平均值的10%以内,而需要大约40个样本才能估计出与平均值的5%以内。
基于上述建议的样本区域数量和每个区域的样本数量,一个全球采样计划,每个样本有3或4个深度和3或4个粒度分数,将需要20万至30万次碳含量分析。这么多样品的分析是不可能的。然而,分层抽样方法提供了一种机制,可以将许多等效的单个样本“散装”为单个“分层”样本,包括局部变异性,并将所需的分析工作量减少到大约12,000个样本。
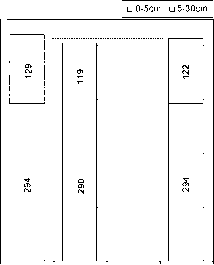
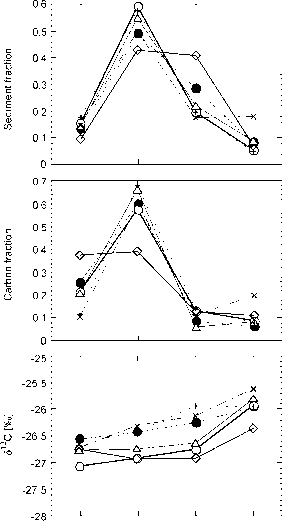
要使这种方法可行,它必须能够再现由来自同一地区的单个样本的许多结果平均而得到的结果。图4显示了从昆士兰北部未放牧的热带稀树草原上玄武岩衍生土壤的两个样带点(约1000米长)采集的单个样本的结果(Bird, 1998)。它还显示了通过对树冠下区域和远离树木区域的这些数据进行平均计算得出的碳储量,以及根据该地点的树冠覆盖百分比加权得出的该地点的平均碳密度。个别样品之间有相当大的分散,但在每个地点平均10个样品,总库存量非常相似(408 mg/cm2 vs 422 mg/cm2 0-30 cm)。
图5提供了这些结果(每个地点40次分析)与使用分层样品方法获得的结果(每个地点4次分析)对同一地区玄武岩土壤和砂质花岗岩来源的土壤的比较。使用每种方法获得的碳清单给出了基本相同的结果,并突出了花岗岩和玄武岩土壤之间碳清单的巨大结构控制差异。单个样本和分层样本方法的结果之间的一致性是可以预期的,因为碳库存之间的混合(即单位体积)可以预期是线性的。
未放牧的玄武岩库存
擦伤花岗岩库存
未放牧的玄武岩库存
HU-8
结合
HU-1 HU-2的组合
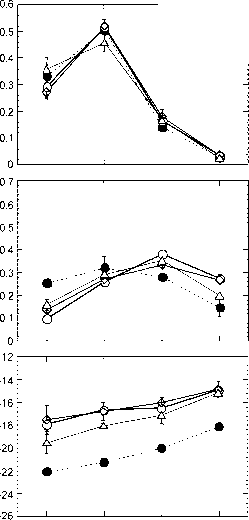
图5图4结果与分层方法的比较,其中a体积分数将每种样品类型的微量元素添加到大样本中,从40个单个样品中生成4个现场平均样品(0-5T, 0-5G, 5-30T和5-30G)。对这些样本进行了单独的分析,对个别样本进行了根据冠盖百分比加权的库存。图4显示了玄武岩源性土壤和砂质花岗岩源性土壤的结果。
0 HU-2(未放牧的粘土)-G
1 HU-1(未擦除粘土)-T 9 HU-2(未擦除粘土)-T
0 50 100 150 200
图6图4所示样品SOC值与土壤碳密度的关系
来自这些样本的S13C结果(图6)支持了以下结论:碳源(树木和草)之间的混合是线性的,因此对分层样本的分析为采样区域提供了具有代表性的值。津巴布韦砂土和粘土的相似结果进一步证实了这种关系(Bird et al, 2000),如图7所示。随着碳密度的增加,S13C值有规律地下降,这与C、树木下的区域单位面积碳含量较高和/或ci衍生碳进入有机碳库后停留时间较长有关。在5 cm深度以下,由于降解对有机碳同位素组成的影响,以及该深度的有机碳与目前覆盖在该地点的植被之间的耦合不太紧密,这种关系不太明显。
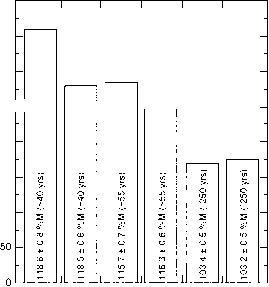
图8显示了来自超过700个单独样本的结果,这些样本被分成六个区域样本,覆盖了加拿大西部的一个大气候梯度。这些结果表明,0- 5cm深度区间碳储量的下降与o的下降一致
公司的CD
公司的CD
图8从美国-加拿大边界到加拿大北极,经过阿尔伯塔省和西北地区的样带上,分层样本0-5厘米区间的碳密度(大约75个样本,每个区域一个样本)。“生物温度”(由Holdridge, 1947定义)在Cypress(50°N)的10.5°C到Windy(68°N)的1.5°C之间,而降水(不包括降雪)在相同的两个地点分别在750到60毫米之间。还显示了63- 500吉特分数的14c活度,这表明表观停留时间从- 40年持续增加到- 250年,同时温度和降水下降。在所有情况下,表面层都被视为“垃圾”层的顶部,数据参考< 2000-/xm分数。
M13(砂)- T |
|
•阿 |
|
图7津巴布韦Matopos研究站的砂土和粘土5”C和有机碳密度之间的类似关系(Bird et al., 2000)。自实验开始(1947-48)以来,这种关系似乎与强加的火灾制度无关,从每年燃烧到完全免于火灾。
导致凋落物厚度和o层逐渐减小。这些数据表明,利用分层抽样方法可以阐明土壤有机碳的区域趋势。
图9比较了不同沙质土壤采用该方法得到的结果气候地区.碳储量从水资源有限的热带稀树草原向两个方向增加潮湿的热带站点和寒冷的高纬度站点。此外,在0 ~ 5 cm区间内,寒冷高纬度站点的总库存量所占的比例高于低纬度站点。这两种观察结果都与前几节讨论的趋势一致,即高纬度土壤中出现厚凋落物和o层,而在高纬度土壤中则没有低纬度的土壤。
这些地点之间的总库存范围约为3倍,但应注意的是,没有潮湿的高纬度地点
450 400 350 300 250 200 150 100 50 0
砂质土壤碳储量□0-5 cm□5-30 cm
休登21”,600毫米
Matopos 18", 630mm
约克角27”,1200毫米
Zotino 4”,300mm
图9使用分层抽样方法产生的结果的比较不同气候条件下raybet雷竞技最新,都在沙质土壤上。在所有情况下,表面层被认为是“窝”层的顶部,并指<2000-/^111分数。雷竞技csgo数据来源如下:Matopos,津巴布韦:Bird et al. (2000);休根登,澳大利亚:鸟(1998);澳大利亚约克角:Kracht和Bird(未发表数据);佐蒂诺,西伯利亚:鸟(未出版。数据)
例如,埃德蒙顿样本的0- 5厘米库存量(图5)与约克角和佐蒂诺地点的0- 30厘米库存量总量大致相同,因此埃德蒙顿地点的0- 30厘米库存量预计会比这两个地点都大得多。
分层抽样方法的另一个优点是,它可以在大大减少的样本集上执行一组更耗时的分析。例如,图10显示了粒度分布0 ~ 5cm间的碳分布和碳同位素组成。尽管两组数据中的矿物粒度分布相似,raybet雷竞技最新气候、植被在不同大小的组分之间,火的作用使碳和碳同位素的分布具有非常不同的特征。同样,这些差异很容易根据前几节中描述的过程来解释(参见图标题)。
分层样品也适用于放射性碳分析,与使用任何现有方法进行特定地点研究相比,分层样品可以为特定气候/土壤质地条件提供更好的“平均”周转时间(例如,Bird等人,1996年;Trumbore, 1993;Harrison et ai, 1995)。63- 500ju的14C结果。图8给出了上述分层加拿大样品的m分数。同样,该尺寸分数中的碳的表观周转时间随着温度和降水的降低而增加,这与基于前几节讨论的预期一致。
使用微生物碳的14c活性(Ladd和Amato, 1988)或微生物呼吸的二氧化碳来获得土壤样品中有机碳平均周转时间的综合测量,这是一种尚未被探索的可能性。这种说法背后的基本原理是,当微生物碳库快速翻转时,微生物碳库的14C活性将由被代谢的底物的量加权14C活性决定。因此,如果顽固的降解碳没有被微生物代谢利用(因此与十年-百年的全球周期无关),它将不会被记录在微生物碳或微生物呼吸CO的14c活性中。
在实验室中,从无根土壤中微生物呼吸的CO的14c活性的测量在技术上是简单的(Santruckova等,出版中),但是存在这样的可能性,即从其田间位置扰动土壤可能使“受保护的”碳可用于代谢。从土壤样品中物理分离微生物碳需要制备两个样品(Jenkinson, 1988)。第一个样品用K2S04提取,第二个样品用氯仿熏蒸后用K2S04提取。氯仿熏蒸使K2S04中有一定比例的微生物碳可萃取物。用质量平衡法测定微生物碳的比例和14c活性。虽然这种方法具有在采集时提供微生物碳的瞬时“快照”的优点(假设在采集后不久通过干燥或冷冻停止微生物活动),但必须进行质量平衡计算,这给微生物14C活性的计算带来了额外的不确定性,而且也不清楚薰蒸步骤是否只释放微生物碳(Badalucco等,1992)。
作为这种方法的初步测试,使用这两种技术分析了代表加拿大样带上“最温暖”(埃德蒙顿)和“最冷”(Windy)位置的样本。图11显示了使用这两种技术获得的结果。微生物呼吸的结果表明,合理的co2周转时间略短于同一土壤的63 ~ 500 × xm部分。用熏蒸萃取技术得到的结果,表观周转时间短,误差大。对两种结果之间的差异的一种解释是,微生物并没有在吸收和呼吸过程中平均分配所有的碳。年轻的富含能量/营养的碳更有可能被分解成生物质,而年老的碳则被用来支持呼吸(J. Schimel,个人交流)。埃德蒙顿样品的微生物碳,4C活性高于1996年预测的14C活性与周转时间之间的关系,这一事实进一步表明,碳原子在该样品中被呼吸或转移到“慢/被动”碳池之前,平均可以在微生物碳池中循环约10年。
这些初步结果表明,与烟熏提取技术相比,呼吸CO技术可能提供更可靠的SOC周转时间估计。在沙质土壤中,碳几乎没有机会得到物理保护,因此上述与样品的物理扰动有关的潜在问题可能是可以避免的。
柏树 |
|
埃德蒙顿 |
|
- e - |
水牛 |
Yellowknlfe |
|
- -+- |
雷的 |
- x - - - - - - - - - - - - |
多风的 |
C1(无火灾) |
|
F1(年消耗) |
|
0 - |
F3(3年燃烧) |
F5 (5yr bum) |
在cd
图10加拿大样带(图8)和Matopos火灾试验(图7)分层样品中碎屑颗粒的尺寸分布以及碳的尺寸分布和同位素组成。虽然大多数样品的碎屑尺寸分布类似于砂状,但相同尺寸分数之间的碳分布存在显著差异。碳均匀地存在于加拿大土壤的粗粒度分数中,但在马托波斯样品的细粒度分数中富集程度不同。在Matopos,碳在大小分数之间的分布受火灾频率的控制,随着火灾频率的增加,更细分数中的碳有规律地增加,导致粗物质的燃烧。加拿大样品中碳的á”C值随着粒径的减小而增加~ Woo,这是由于陆地Seuss效应和与之相关的分馏作用微生物降解(Bird and Pousai, 1997)。Matopos样品中碳的5,3c值随着粒度的减小而增加2-3%,这是由于c4衍生碳输入到细组分的差异或C衍生碳在粗组分中的选择性保存(Bird和Pousai, 1997)。加拿大样品的SUC值是典型的高纬度C′森林(Bird et ill。, 1996),而Matopos值表明c4衍生碳的输入是可变的。应当指出的是,Matopos结果是通过在每种情况下遭受每种燃烧制度的两个地点重复分层采样获得的。这些独立重复抽样结果的可比性(由误差条表示)进一步证明分层抽样方法可以产生可靠的结果。
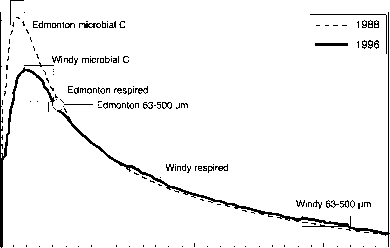
图11微生物呼吸co2、微生物碳(熏蒸-萃取)的14C活性和63- ~ 500-/u。图8还显示了停留时间与4C活性之间的关系,1996年收集样品时预计4C活性与1988年的4C活性之间的关系,1988年是上次在有机碳池中观测到的Windy微生物碳样品中如此高的值。
居住时间[年]
图11微生物呼吸co2、微生物碳(熏蒸-萃取)的14C活性和63- ~ 500-/u。图8还显示了停留时间与4C活性之间的关系,1996年收集样品时预计4C活性与1988年的4C活性之间的关系,1988年是上次在有机碳池中观测到的Windy微生物碳样品中如此高的值。
6.结论:《沙世界》和《粘土世界》
前几节的讨论已经表明,SOC库中观察到的趋势背后的机制已经被理解,并且已经开发了模型,可以充分描述在记录良好的现场站点观察到的碳分布模式。进一步的讨论还表明,在采样和分析协议方面具有一致性,足以为全球模型提供进一步约束的此类站点数量很少。
虽然它并不打算取代在个别地点进一步的面向过程的研究,但有人认为需要一种新的SOC数据类别,旨在细化我们对SOC行为的决定因素如何在全球范围内相互作用的理解。这种类型的数据必须使用来自全球分布地点的一致方法,以产生关于有机碳储量、通量、潜在呼吸和同位素的信息。这些信息必须始终与纹理和深度分布数据相耦合,这些数据考虑到当地的可变性,并能够以适合与模型派生数据比较的格式提供区域规模的估计。在这个方向上已经做出了一些努力(Paustian et al., 1995;Falloon等人,1998)。
沙世界和粘土世界,即从分布在全球气候带的整个光谱的粗和细纹理基材中对SOC参数的观测估计,在技术和后勤上是可以实现的。它们代表了基于过程的特定地点研究的自然延伸,将需要进一步细化我们对全球范围内SOC池的理解。要解决当前围绕全球碳循环运作的主要问题,并回答因批准《京都议定书》而产生的政治问题,就需要这样一种改进。
鸣谢
马克斯·普朗克生物地球化学研究所提供的访问奖学金极大地促进了这篇手稿的完成。
参考文献
艾特杰,克特纳,L. P.,杜文诺,P.(1979)。陆地初级生产和植群。在“全球碳循环范围”中。(B. Bolin, E. T. Degens, S. Kemper, and P. Ketner, Eds.),第129-181页。威利,纽约。卷。13。葛兰,g.i,布萨塔,E.和巴勒登特,J.(1996)。有机质分解过程中的同位素鉴别:理论分析。土壤科学。点。 J. 60, 1121-1126. Archer, S. (1990). Development and stability of grass/woody mosaics in a subtropical savanna parkland, Texas, U.S.A. /. Biogeogr. 17, 453-462. B
继续阅读:植物化合物及其作为土壤有机质的转化与稳定
这篇文章有用吗?
读者的问题
-
比尔博罗珀3个月前
- 回复