木本植物入侵和生态系统生物地球化学
土壤和小气候的改变伴随长期过度放牧可能转移资产支持Nraybet雷竞技最新2-fixing或常绿木本植物比草更好适应贫瘠土壤和温暖,干燥的微环境。乔木和灌木的建立将进一步增强grazing-induced减少草本竞争和火(弓箭手,1995)。此外,拉来一小杯非常不快的木本植物,和浏览由野生动物或牛是最小的。然而,水果占主导地位的树的入侵者(Prosopis glandulosa)很容易被牲畜,分散大量可行的种子到草原(布朗和弓箭手,1987)。因此,沉重的,连续的,和优惠的草放牧牲畜促进了木本植物入侵通过大量的直接和间接影响(弓箭手,1994)。作为放牧草原伍迪社区发展,植物和土壤C和N池大小和通量率变化在以下部分中描述。
3.3.1植物碳池
定量变化木本植物覆盖在拉来一小杯描绘在图4和图5。确定这些植被变化对植物碳储量的影响,我们与世纪植物演替模型拉来一小杯(斯坎兰和弓箭手,1991)。我们发起的木本植物入侵在1800年代末严重擦伤了,免费的格局,SOC的内容被放牧降低(图3),景观组成的
65 r
- b - 1 - 2 #■-景观景观…问景观3
图4总木本植物覆盖的变化在三个复制在德克萨斯州南部风景在洛杉矶来一小杯。参见图5的空间格局变化各种补丁类型在景观1(阿切尔Boutton,未发表)。
- b - 1 - 2 #■-景观景观…问景观3

图4总木本植物覆盖的变化在三个复制在德克萨斯州南部风景在洛杉矶来一小杯。参见图5的空间格局变化各种补丁类型在景观1(阿切尔Boutton,未发表)。
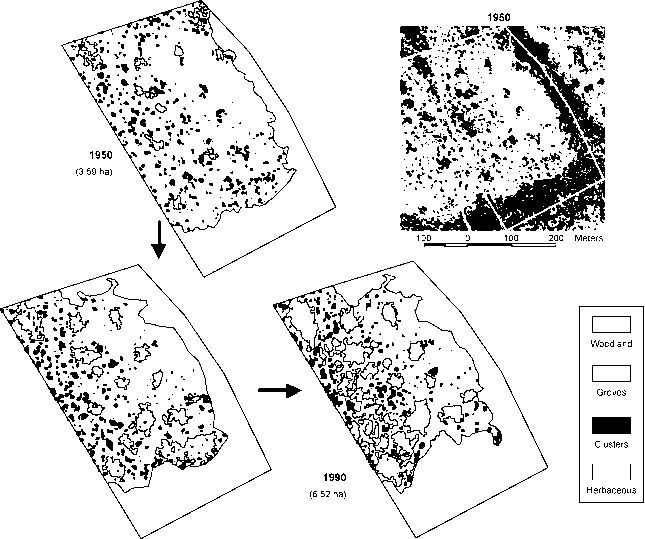
1976年
图5景观尺度草本和木本植物群落覆盖的变化从1950年到1990年在高地(草本、离散集群和林)和低地(林地)植物群落在德克萨斯州南部的洛杉矶来一小杯(阿切尔Boutton,未发表)。值下面日期公顷的木本封面(grove集群+ +林地)11.06公顷的“像素”。See Figure 4 (Landscape 1) for changes in percentage woody cover.
1976年
图5景观尺度草本和木本植物群落覆盖的变化从1950年到1990年在高地(草本、离散集群和林)和低地(林地)植物群落在德克萨斯州南部的洛杉矶来一小杯(阿切尔Boutton,未发表)。值下面日期公顷的木本封面(grove集群+ +林地)11.06公顷的“像素”。See Figure 4 (Landscape 1) for changes in percentage woody cover.
砂质壤土高地和粘壤土间歇排水、木本植物填充基于rainfall-driven过渡概率。草原和森林世纪应用子程序,适当的,计算植物碳和SOC(0-20厘米)的森林和草本股市景观元素。在每个时间步,植物碳和SOC估计总结整个景观(高地和低地植被斑块类型)。结果表明,当今的发展草原parkland-woodland复杂植物碳储量增加了10倍,这将是目前的“原始”草原植被一直保持在网站上(图6)。增加的部分原因是由于地上净pri玛丽生产力的增加(表2)和它的一部分代表组织营业额的下降,取而代之的是木本植物草本植被时发生。这些结果保守在世纪模拟包括根C质量只有在前20厘米的土壤。木本植物根系的生物量分布在拉来一小杯(美国瓦茨,1993;Boutton等,1998;1999;切割Midwood等,1998;吉尔和伯克,1999)是典型的其他旱地树/灌木系统(杰克逊等人,1996; Canadell et al, 1996) where, relative to grasslands, there is substantially greater mass at deeper depths where turnover and decomposition are likely to be reduced. The fact that fluctuations in monthly woody plant root biomass in upper soil horizons exceeded monthly foliar litter inputs by one to
表2对比在地上净初级生产(友好),土壤物理性质、有机碳和总氮池(0 - 10厘米深),和通量在土壤与木本植物和放牧草地群落在砂壤土旱地景观拉来一小杯在德克萨斯州南部研究领域
社区类型
参数
草本
表2对比在地上净初级生产(友好),土壤物理性质、有机碳和总氮池(0 - 10厘米深),和通量在土壤与木本植物和放牧草地群落在砂壤土旱地景观拉来一小杯在德克萨斯州南部研究领域
参数
草本
木本植物
进行友好(Mgha-1第一年)” |
1.9 |
-3.4 |
5.1 |
- 6.0 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
体积密度(g厘米”!)J |
1.4 |
±0.01 |
1.1 |
±0.04 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
%粘土的 |
20. |
±0.7 |
20. |
±1 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
细根(g m ~:) J |
One hundred. |
-175年 |
400 - |
- 700 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
粗根(g m - 2)”1 |
One hundred. |
- 400 |
400 - |
- 1100 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
有机C ' % |
0.84 |
- 0.05 |
2.2 |
±0.23 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
gni-2 |
1165年 |
±67 |
2352年 |
±276 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
潜在的C矿化 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(毫克公斤“土壤普通人)1 |
7.3 |
±5.7 |
15.5 |
±6.8 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
土壤呼吸(mg公司;m - 2第一年)” |
611年 |
±83 |
730年 |
±67 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
瞿)值原位土壤respiration1。 |
1.2 |
1.4 - |
- 2.7 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
总N ' % |
0.07 |
±0.00 |
0.18 |
±0.02 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
gm-2 |
91年 |
±6 |
192年 |
±20 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
N矿化”1,g N m 2 1年 |
6 |
±1 |
22 |
±2 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
MgNg-1第一年 |
42 |
±5 |
200年 |
±18 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
没有通量(ng没有n cm-2 h - !)” |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
干土 |
0.2 |
±0.07 |
2.8 |
±0.25 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
湿润的土壤 |
1.1 |
±0.11 |
16.2 |
“原始”草原 ——严重擦伤了草原 我|草+伍迪补丁 木本植物 请注意。每月最大和最小值获得的样品在一个年度周期显示root-standing作物(粗根> 0.1毫米直径);提出了一系列友好和Q, 0。其他值都意味着±SE。 希巴德,1995。McCulley, 1998。“科尔et al . 1996。 两个数量级(表2)表明地下的有机质的输入驱动土壤理化性质变化后续木本植物建立草原。这些木本植物根生物量大幅波动表明高营业额,这是符合详细观察木本植物在其他系统(Eissenstat柳井正,1997;亨德瑞et al ., 1997)。此外,草根周转率可能低于普遍认为(Milchunas et al ., 1992)。因此,增加地上和地下的净初级生产力可能伴随木本植物侵入草原和促进C和N的积累。 作为一个独立的测试重建在图6中,我们量化地上植物碳密度在补丁代表占主导地位的社区类型来一小杯。这是完成了使用异速生长的关系和样带调查(诺萨普et al ., 1996)。植物碳密度然后乘以社区面积测量航拍照片(1950、1976、1990)获得社区水平的估计。估计每个社区类型然后总结获得景观尺度的估计。对于补丁代表各种木本和草本群落类型,世纪的估计地上碳密度低于实地评估(表3),进一步表明模型估计是保守的。地上碳之间的密度差异显著 1900 1910 1920 1930 1940 1950 1960 1970 1980 1993年 图6模型的改变真个场面(高地+低地的气息,在其中所有补丁类型)植物碳密度(地上+根20厘米)陪同继承从草原到草原公园/林地(从希巴德,1995)。虚线描述稳态SOC预期的轻轻掠过草原景观(高地+低地社区汇集)每10年用火,没有木本植物;实线描述稳态植物碳密度很大,连续放牧草地景观没有火,没有木本植物。木本植物丰度的变化在每个土壤类型是由一系列模型(斯坎兰和弓箭手,1991);然后评估后续植物碳储量的变化和生物地球化学模型(世纪;Par-ton胡麻。,1994)。为验证结果见图7。 三个景观理清了1950年(图7),主要反映不同木本植物覆盖在这个日期(图4)。到1990年,伍迪封面和碳密度可比在三个景观。CENTURY-succession模型估计地上碳密度的“平均”景观,1950年,1976年和1990年密切近似从field-historical天线获得这些照片的方法。 3.3.2。Nonmethane烃通量在区域的基础上,从草地上转移到木本植物统治有潜在影响的生物物理方面land-atmosphere交互,如反照率、蒸散、边界层条件,和尘埃加载(例如,科比et al ., 表3观察和预测地上碳密度在补丁代表树/灌木和草地社区来一小杯,德克萨斯州 碳密度(公斤iyT 表3观察和预测地上碳密度在补丁代表树/灌木和草地社区来一小杯,德克萨斯州 碳密度(公斤iyT
请注意。观测数据(意味着±SE)是基于带横断面和木本植物size-biomass关系社区(阿切尔Boutton,未发表),在剪块草原(阿切尔未发表)。世纪预计值估计为100岁高龄的补丁(希巴德,1995)。 的峰值地上部生物量。 景观:我二三世II三世II三世 1950 1976 1990 图7在地上植物碳密度变化在三个景观从1950年到1990年在洛杉矶来一小杯(阿切尔Boutton,未发表)。补丁/ soil-specific场估计的植物碳密度(马克穆特诺萨普et al ., 1996年,尼尔森和弓箭手,未发表)乘以贴片面积来衡量在航拍照片生成真个场面估计。虚线表示从链接CENTURY-succession模型预测(希巴德,1995)。参见图4为伍迪盖三个景观的变化。 1990;Pilke Avissar, 1990;格雷茨,1991)。植被的变化也可能影响大气氧化能力,气溶胶的负担辐射特性影响non-methane碳氢化合物的排放(NMFlCs)如萜烯、异戊二烯和其他芳烃(Fehsenfeld et al ., 1992)。有许多来源的大气NMHCs但> 90%的全球年度排放来自植被(Guenther et al ., 1995)。因此NMHC的排放高度依赖物种组成是受到环境条件影响植物生理学和生产。高温度和太阳辐射通量与亚热带和热带草原和热带稀树草原使这些地理上广泛的生物气候地区大型生物的潜在来源NMHC的排放。然而,草通常NMHCs排放低,而排放的树木和灌木森林系统是高度可变的,一些物种是低排放和其他物种被高排放者。 我们假设在木本植物叶面NMHCs排放与叶呈正相关长寿光合能力和成反比关系。植物的特点是低的光合能力和缓慢增长利率(如常青树)取决于延长叶片寿命达到积极的碳平衡。优先分配次生化合物如萜烯将帮助确保叶寿命减少的食草性水平。物种较低的光合能力和高水平的次生化合物也应该主导下木排除高增长率低光照水平的环境中,植物更容易浏览器。相比之下,物种选择竞争力光合率高,高增长率和高营业额的组织(例如,落叶灌木)。分配到二级化合物阻止食草性的低优先级叶寿命不是实现一个积极的叶面投资回报的关键。这种植物会优先分配资源,如氮羟酶和富有成效的组织而不是结构组织或次生化合物如萜烯,并将因此低NMHC的排放(或异戊二烯的排放,因为异戊二烯是不清楚与国防有关;Coley et al ., 1985)。 要测试这些假设,我们筛选植物物种代表的主要增长形式在拉来一小杯NMHC的排放。正如所料,草NMHC的排放较低的利率和几种常见木本种类有高发射率(Guenther et al ., 1999)。Flowever,几乎没有证据表明持续排放的木本植物分类、发展形式,或官能团。结果,概括关于NMHC的排放光谱树/灌木物种组合在其他系统中没有出现可行。 确定生物NMHC的排放已被改变的土地覆盖的变化从草、木本植物统治在洛杉矶来一小杯,植被变化模型(斯坎兰和弓箭手,1991)是与一个模型预测NMHC的排放作为叶面密度的函数,叶片温度,以及光合光量子通量密度调制到环境温度,降水、相对湿度、云量风速(Guenther et al ., 1995;尔德1997)。 生物排放模型的链接与植物演替模型表明,自1800年代初以来土地覆盖变化引发异戊二烯的排放增加三倍(图8)。这一增长反映了植被组成和增加叶面密度的变化。模型预测当前NMHC的排放中有20%的塔的通量测量系统。详细的实地测量两种常见灌木物种表明,异戊二烯的排放与叶片温度的增加呈指数增加从20到40°C和没有抑制干旱压力。因此,模型预测,预计2 x-c02气候下,当今生物NMHC的排放将增长一倍。raybet雷竞技最新 这些估计的变化与转换关联NMHC的排放的草原林地依照估计其他生态系统。例如,科林格et al。(1998)记录增加四倍总萜类化合物排放单位叶片质量以及林地横断面在中部非洲的稀树大草原。这些变化与植被变化有关NMHC的排放在德克萨斯州的亚热带和热带非洲也反映报道对温带森林(马丁和尔德1995)。这些结果显示的大小NMFIC排放时可能发生的变化raybet雷竞技最新气候和植被作文是改变。这些增加的重要性NMHC的排放是放大在拉来一小杯,发生与升高一氧化氮(NO)排放shrub-modified土壤(表2;3.3.4看到秒。细化)。 为什么vegetation-induced增加NMHC的关注?生物碳氢化合物发挥重要作用产生污染物如0(公司,和有机过氧化物,而影响氢氧自由基(OH)减少大气化学oxida - 历史(1800)草原/稀树大草原 建模= 0.72 = 0.54 今天草原公园/刺林地 土地覆盖变化t放牧4。浏览4火 异戊二烯通量(mg 2 h_1 C m”) 测量建模 建模= 0.72 = 0.54  测量建模  图8的变化nonmethane烃(异戊二烯)排放预测陪从草原草地转向稀树大草原林地在德克萨斯州南部的洛杉矶来一小杯(基于Guenther et al ., 1999)。从耦合succession-NMHC发射模型预测与测量值相比于通量塔。所示的“测量”价值观的历史景观从塔位于草原草地景观伍迪覆盖较低。 变形能力和增加温室气体的停留时间。据估计,来满足当前空气质量标准,对流层臭氧、人为碳氢化合物的排放量会减少只有30%没有天然的异戊二烯的排放,但其中70%的存在(曼森et al ., 1991)。NMHC-NO排放量的变化与区域转换的草原灌丛带可能因此构成的“基线”测量对流层臭氧生产由汽车尾气排放和工业来源。 3.3.3土壤C和N池连接建立后,木本植物改变它们周围的土壤和小气候影响池大小和通量的营养素。raybet雷竞技最新结果是形成“生育的岛屿”,这一现象被广泛量化旱地(见查理和西部,1975;施莱辛格et al ., 1990;斯科尔斯和弓箭手,1997;特殊生物地球化学问题42 (1/2)1998)。三个通用机制提出了占这个(举例来说,维吉尼亚州,1986):(1)木本植物作为营养泵,画深层土壤的养分视野,从树冠之外地区的外侧,存放他们林冠下通过茎流、litterfall,树冠浸出;(2)高,空气动力学的木本植物的树冠陷阱nutrient-laclen大气尘埃,雨洗掉叶子和subcanopy土壤;和(3)木本植物可以作为焦点吸引栖息鸟类、昆虫和哺乳动物寻找食物,阴影,或覆盖。这些动物可以通过排便和挖掘土壤的肥力。由于这些原因,土壤碳和氮池应该增加后续在放牧草原木本植物殖民。 在拉来一小杯,地表(清廉厘米)土壤与木本植物已知了过去一个世纪有较低的体积密度,含有更多的根生物量、更高浓度的SOC和总N,并有呼吸和N矿化率大于土壤与其余放牧草地群落(表2)。随着木本植物覆盖的连续性增加通过时间,景观尺度土壤养分池和通量预计将增加,变得更加均匀分布。因此,有关CENTURY-succession模型运动(见3.3.1节)预测,到1950年,景观尺度SOC水平回到这将发生的“原始”草原被保持在网站上(图9)。在1990年代早期,景观尺度SOC含量约10%高于预期的“原始”草原,和大约30%高于大量放牧草地没有经历木本植物入侵。模型预测SOC沉积将持续数百年,达到均衡水平三倍的今天的放牧草地的社区。 虽然生育的“岛”现象已被广泛认可,所知甚少的养分富集率的tree-dominated补丁。总C和N在土壤金合欢塞内加尔和Balanites aegyptiaca树冠呈正相关,与树周长(r2 = 0.62和0.71,分别;Bernhard-Reversat, 1982),表明木本植物的净积累时间占用一个补丁。在温带老油田进行森林演替土壤碳储存增加了40%在植物+池超过40年(约翰斯顿等,1996)。在拉来一小杯、土壤C和N下量化Prosopis glandulosa树木的年龄是由年轮数决定。土壤有机碳存储(前20厘米的土壤)线性树茎年龄增加
1900 1910 1920 1930 1940 1950 1960 1970年 1900 1910 1920 1930 1940 1950 1960 1970年y 1980年 1993年 图9的变化土壤有机碳(SOC);0-20厘米)预测伴随木本植物侵入放牧景观组成的砂质壤土高地和粘壤土间歇性流域拉来一小杯网站(从希巴德,1995)。虚线描述稳态SOC预期的轻轻掠过草原景观(高地+低地社区汇集)每10年用火,没有木本植物;实线描述了大量的稳态SOC,不断放牧草地景观没有火,没有木本植物(见图3)。酒吧表示SOC总结整个景观和包括草原和木本植物群落。注意,到1950年,SOC水平增加了水平可比的“原始”草原(交叉线棒)。木本植物丰度的变化在每个土壤类型是由一系列模型(斯坎兰和弓箭手,1991);后续的变化然后评估土壤碳和生物地球化学模型(世纪;帕顿等< / /。, 1994)。 率从11.8到21.5 g C m 2第一年在砂壤土高地伍迪补丁类型47.2 g C m - 2第一年所,粘壤土林地斑块(表4),利率总N的积累(前20厘米的土壤)范围从1.9到2.7 g N m - 2第一年在桑迪高地和平均为4.6 g N m - 2第一年在粘壤土低地。然而,木本植物年龄解释只有21 - 68%的土壤C和N的变化削减利率。这些低r2值可能表明树茎年龄不准确地反映植物的年龄,可能是因为过去的干扰和后续营养伍迪封面的再生。低r2也可能表明,联合国相关因素树的入住率影响土壤C和N下树的树冠。这些因素可能包括小规模的异质性与大型哺乳动物或鸟类排便,土壤混合小型哺乳动物和节肢动物或patch-specific物种组成差异,生产力和发展速度的林下叶层社区。 建模实验允许我们控制的因素可能会导致实地的变化估计木本植物age-SOC关系。模型估计的SOC积累与领域高地补丁类型和估计大大低于字段低地估计补丁类型(表4)。模型估算土壤N积累显著低于估计领域,特别是在低地。鉴于伍迪补丁年龄只有26 - 68%的方差解释土壤C、N含量,我们这个领域累积利率不能作为最终的估计。模型低估了野外观察结果,特别是对n的可靠性模型估算土壤碳可以用更好的理解可能会提高营业额的大幅根质量(表2)可能会在补丁类型之间存在差异。模型估算土壤N可能受制于缺乏与N2固定相关信息输入,大气N沉积,高地和低地之间的易位,根营业额。 3.3.4土壤C和N动态C和N池的增加土壤与木本植物群落发展中放牧草原上拉来一小杯都伴随着土壤呼吸增加,N矿化,一氧化氮(NO)排放(表2)。没有通量的增加伴随扩张的木本植物在草原上拉来一小杯是值得注意的。一氧化氮在大气化学中扮演几个关键作用导致酸雨和催化的形成光化学烟雾和对流层臭氧。后者可能是强调在洛杉矶来一小杯设置,因为没有和碳氢化合物的排放(见秒。3.3.2)与此同时升高之后木本植物。 有机质的数量和质量的输入交互驱动土壤代谢活动(Zak et al ., 1994)。因此,年度土壤
请注意。字段数据来自补丁年龄(取决于之间的线性相关性树木年代学)和土壤C和N质量(Boutton和弓箭手,未发表)。从世纪模拟模型估计(希巴德,1995)。的描述对比伍迪补丁类型可以在阿切尔(1995 b)。 请注意。字段数据来自补丁年龄之间的线性相关性(由树木年代学)和土壤C和N质量(Boutton和弓箭手,未发表)。从世纪模拟模型估计(希巴德,1995)。的描述对比伍迪补丁类型可以在阿切尔(1995 b)。 呼吸率和净初级生产力呈正相关(赖斯和施莱辛格,1992)。高碳通量观测的发展木本社区在半干旱La来一小杯草原可能反映了根(表2)和叶生物量增加木本植物的树冠下的输入和增强土壤水分(通过集中降雨从茎流、液压升降机、和/或减少蒸发)。在一起,这些生物和非生物因素相互作用来刺激微生物活性相对于grass-dominated土壤。事实上,微生物生物量在伍迪社区相当或高于草原社区在拉来一小杯(McCulley, 1998)。然而,实验灌溉,缓解植物水的压力、增强光合作用(马克穆特,1997)和土壤呼吸增加,引起土壤微生物生物量下降。这表明,升高在木本植物群落土壤呼吸观察到洛杉矶来一小杯可能会因此更加根生物量的变化(表2)和呼吸比微生物生物量的变化和活动。 来估计土壤景观尺度变化有限公司通量,我们增加补丁/ soil-specific估计年度土壤呼吸(McCulley 1998)补丁。然后我们计算贴片面积的变化与连续模型(斯坎兰和弓箭手,1991)。景观尺度土壤呼吸(公斤C是第一年)预计将增加从6687 (200 YBP)到7377(1990年代)到7602(200年以后)(表5)。这增加了10.3%从历史性的草原草原过渡到现在的稀树大草原parkland-thorn林地复杂,增加一个额外的3%发生如果目前草原森林公园的发展。如果年平均气温增加投射在环流模型,进一步增加土壤呼吸将(所有其他因素不变的情况下)。的确,在木本植物群落土壤呼吸的Q10值(1.4、2.7和2.3集群,树林,和林地类型,分别)超过放牧草原(1.2)在拉来一小杯(McCulley, 1998)。这表明,如果未来温度变化发生,最近的重要性和预计未来植被变化对土壤呼吸将进一步放大。例如,增加土壤呼吸的大小从过去的草原草原垫为22.4°C到未来林地垫为28.4°C会是22.5% (C = 6687 - 8197公斤是第一年;基于赖斯和施莱辛格,1992)到99.3% (C = 6687 - 13328公斤是第一年;基于McCulley, 1998)(表5)。潜在的变化量,季节性,和有效性的降雨将有着重要的、但还未可知,对这些预测的影响。 3.3.5土壤源和汇高通量的C和N从植物和土壤grass-land-to-woodland转换后在拉来一小杯表明潜力增加温室气体的积累和改变tropos-pheric化学,尤其是木本植物入侵已在地理上广泛(如表1)提出的。然而,正如秒。3.3.3,土壤有机C和N积累了发展木本植物群落在拉来一小杯,尽管高通量和高流动率。这表明输入超过输出,土壤和植被在拉 表5预计景观尺度年度土壤呼吸的变化(SR;公斤C农业年”)陪同继承从打开的稀树大草原/草地林地和潜在的年平均温度的变化 景观尺度土壤呼吸(公斤C是第一年) 未来的刺 年平均过去草原现在草原林地 温度(°C) (200 YBP)”1公园/林地复杂(200 YAP) 1, 答:基于垫/地图回归在赖斯和施莱辛格(1992) 22.4 6687 7377 7602 25.4 6948 7666 7899 28.4 7209 7954 8197 b .基于Q,从McCulley„值(1998) 25.4 8083 9938 10465 28.4 9480 12499 13328 请注意。(补丁)-草和各种伍迪社区和老soil-specific率测量月度/年度周期在拉来一小杯(McCulley, 1998)的面积乘以各自的社区类型(斯坎兰和弓箭手,1991 j。年平均温度变化的影响(SR垫、°C)从(A)估计方程在赖斯和施莱辛格(1992);拉来一小杯(垫- 22.4°C和地图- 720毫米)3和6°C增加垫会产生增加7.8%和3.9,分别在土壤呼吸;和(B) Ql0原位的值,会提供对基层社区适用的土壤呼吸McCulley (1998)。可能是保守的估计,随着呼吸率低于正常降雨期间用于计算测量。“YBP,年现在之前。h狂吠,年后礼物。 来一小杯功能,C和N汇过去一个世纪。多种因素可能占观察交互增加土壤C和N池: •流离失所的树木和灌木草可能更有效率abovcground和bclowground因此交付更多的有机物质进入土壤根生物量和友好(见表2)。 •1、屋檐豆科和豆科木本植物在拉来一小杯[N]高于草(2 - 4% vs < 1%;阿切尔未发表)。然而,木本植物在这些风景弧很少浏览通过家畜或野生动物,表明高浓度的次生化合物。这可能导致一个重大垃圾质量X数量互动,即 ••叶生物量的大部分由树木和灌木直接进入土壤池和垃圾而不是通过食草性途径,和••叶从木本植物生物质输入一个更大的部分可能是耐分解。 •伍迪垃圾输入和粗,木质化的灌木的根将促进C和N积累比草“根与芽”。 •阴影/灌木树的树冠减少土壤温度相对于那些在草原(弓箭手,1995 b),因此限制潜在的矿化(Q, 0效果)。 •氮积累的结果是一个潜在的N2固定网站通过豆科灌木常见(p . glan-dulosa和几个金合欢spp。)和/或N的吸收和横向易位草原补丁。尽管nodula-tion诱导在控制环境和现场条件下观察到洛杉矶来一小杯网站(Zitzer等,1996)和其他地方(弗吉尼亚等人工智能,1986;约翰逊和麦克斯,1990),方法论的限制阻止了N2固定量化(·汉德里Scrim-geour, 1997;廖et al ., 1999)。根分布研究(美国瓦茨,1993)折扣外侧觅食的假设。 •拉来一小杯内ca。70公里的一个主要炼油厂中心(科珀斯克里斯蒂,TX)和大气N沉积可能是重要的过去50 - 75年(例如,荷兰et al ., 1999)。N可用性的增加可能促进了木本植物扩张(如Kochy, 1999)通过减轻grass-woody植物对土壤N的竞争和促进经济增长的木本植物多擦过的草。反过来,这可能会转化为更大的有机C和N输入土壤与木本植物有关。 3.3.6一个不确定的未来Prosopis glandalosa目前占据了上层在高地和低地的木本植物群落。根据补丁类型,它构成了40 - 90%的地上生物量(阿切尔Boutton,未发表)和30 - 70%的粗根生物量(> l-mm直径)(美国瓦茨,1993)。因此,p .格兰dalosa必须的动态变化的主要驱动在植物和土壤C和N股在拉来一小杯。未来增加景观营养池和通量将反映的组合(a)持续增长的p . glandulosa和相关的灌木在现有木本植物群落和(b)扩张的木本植物到剩下的草原。 持续扩张的可能性有多少?这可能取决于土地管理实践。可以使放松的放牧压力草生物量积累和消防规定(或自然)发生。在一起,这可能妨碍扩张和木本植物的生长。然而,洛杉矶来一小杯似乎跨越了一个阈值,即土壤、种子银行,和植物再生特性,减少放牧压力可能的后果很小(弓箭手,1996)。在草原放牧放松会影响木本植物建立主要通过其影响消防制度(弓箭手,1995;布朗和弓箭手,1999)。然而,剩下的草本空地都很小,间断分布。因此,即使好燃料积累,大火将高度本地化。这种火灾可能防止未来入侵到剩余的草原空地,但不可能把草原木本植物群落,由于树木和灌木在拉来一小杯迅速再生,发芽后干扰(斯坎兰,1988;Flinn等,1992)。费用结算的木本植物,通过机械或化学治疗是禁止的,一般不划算,特别是治疗效果是相对短暂的。 Thus, the likelihood of continued woody plant dominance is high, even with aggressive land management practices which might favor grasses. 木本植物的演替模型模拟了扩张到剩余的草原(斯坎兰和弓箭手,1991)的项目与过度放牧和没有火,伍迪封面将继续增长,直到景观趋于接近完成郁闭。这种假设已经被标明扩展字段数据证实横向根超出伍迪的树冠很小(美国瓦茨,1993)。因此,很少有机会大类间根竞争和密度制约的监管。因此,树/灌木密度可能继续增加,直到所有可用的草本空地已经占领,树冠覆盖几乎是连续的。因此,伍迪补丁对比旱地土壤和伍迪补丁上高地,边境伍迪社区低地种植和合并从1940年代到1990年代(弓箭手等,1988;斯托克斯,1999)。然而,最近的研究表明,拉来一小杯风景可能达到对木本植物的承载能力,由于topoedaphic约束(Stroh, 1995;斯托克斯,1999)。如果是这样的话,未来的变化只发生在C和N池与发展现有木本植物的社区。只有时间会证明这确实是如此。 当前人口占统治地位的p . glandulosa年龄、生长和生物量累积利率应缓慢,除非其他伍迪物种补偿。殖民的林下灌木Prosopis林冠下成立之后在草原Prosopis增长缓慢和种子生产,加速其死亡率(巴恩斯和弓箭手,1998),防止其重建(弓箭手,1995 b)。因此,p . glandulosa似乎不会是未来的一个组成部分林地景观。评估迄今为止表明,从短期来看,损失Prosopis不会影响林下灌木生产力或土壤C和N池(希巴德,1995;巴恩斯和弓箭手,1996)。然而,没有相关的伍迪物种似乎遗传潜力实现成熟Prosopis植物的大小,无论高于或地下。因此,可能会有更少的碳储存潜力一旦失去了从系统Prosopis,除非剩余的林下叶层物种补偿通过提高他们的生产力。此外,目前碳存储在Prosopis生物量将通过死亡和失去分解,虽然比较缓慢。这将是有趣的探索与链CENTURY-succession这些场景模型。不幸的是,我们知之甚少的生产力林下灌木。 Further, the maximum age of P. glandulosa is unknown and we have little basis on which to prescribe mortality from the present-day population. |
继续阅读:养分矿化和植物营养吸收
这篇文章有用吗?


