科琳Le Quere和尼古拉斯Metzl
自然CO2通量
海洋拥有50倍比大气碳。海洋的大型碳含量结果碳化学。当二氧化碳在海洋里溶解,它与水和碳酸盐岩(CO32 -)反应形成碳酸氢盐离子(HCO3 -)根据下列平衡反应:
二氧化碳+水+ CO32 - ^ 2 hco3 - (1)
二氧化碳的总和,HCO3 -和CO32 -被称为溶解无机碳(DIC)。百分之九十一的DIC HCO3 -的形式,CO32 - 8%,二氧化碳的1%。只有二氧化碳可以交换与大气。
在大规模的,现在好了,海洋占二氧化碳主要集中在温带和高纬度地区,而海洋除气二氧化碳通常在热带地区(厚的实线,图12.1)。海气界面二氧化碳通量分布是由二氧化碳溶解度和生物和物理过程。
二氧化碳溶于冷比在温暖的水。当水加热,可以排除的二氧化碳到大气中。这个过程主要发生在赤道地区,上升流带来从深海冷水接触大气(虚线,图12.1)。相反,在中纬度溶解度的效果产生二氧化碳吸收,电流从低纬度地区运输温暖的表层海水。大多数的中间值和深水海洋上一次接触高纬度地区的气氛。因此,二氧化碳在深度平衡在寒冷的气温中,与大气中形成的碳浓度5%,深度比表面(Murnane et al . 1999年)。这个过程叫做溶解度泵。
海洋浮游生物碳和营养。当他们死去或排泄废物,
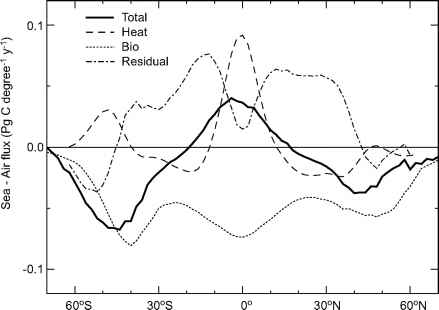
图12.1。海洋空气二氧化碳通量的纬度带(包括学位”1 y-1)。负值表示通量海洋从大气中。总二氧化碳通量估计高桥et al。(2002)根据海洋二氧化碳分压的观察。生物生产出口估计的贡献SeaWiFS叶绿素观测平均在1997 - 2001年,Behrenfeld和法尔(1997)的主要生产、和法律et al。(2000) ef-ratio。热通量的贡献估计使用NCEP热通量平均在1985 - 1995年制定Murnane et al。(1999)。生物学和热贡献与5°平滑运行意味着对应使用的网格高桥et al . (2002)。从生物学和热通量贡献承担无限气体交换。剩余磁通估计从总-生物和热量的贡献。它代表了海洋运输和人为的吸收以及与其他相关的误差估计。
图12.1。海洋空气二氧化碳通量的纬度带(包括学位”1 y-1)。负值表示通量海洋从大气中。总二氧化碳通量估计高桥et al。(2002)根据海洋二氧化碳分压的观察。生物出口生产的贡献估计使用SeaWiFS叶绿素观测平均在1997 - 2001年,Behrenfeld和法尔(1997)的主要生产、和法律et al。(2000) ef-ratio。热通量的贡献估计使用NCEP热通量平均在1985 - 1995年制定Murnane et al。(1999)。生物学和热贡献与5°平滑运行意味着对应使用的网格高桥et al . (2002)。从生物学和热通量贡献承担无限气体交换。剩余磁通估计从总-生物和热量的贡献。它代表了海洋运输和人为的吸收以及与其他相关的误差估计。
死组织要么留在水溶解有机碳(DOC),由电流,运输或聚合成颗粒有机碳(POC),碳汇和乘火车,因为它下跌(虚线,图12.1)。医生和POC remineralized回到DIC生物体和紫外线退化。沉没POC和文档创建一个碳通量从表面到深海,从大气中孤立的地方几十年几百年。单向的碳通量表面深度叫做出口生产。深水,富含碳,回到表面的物理过程(水流和混合)。整个碳的循环出口生产平衡的物理传输介质组成的被称为“生物泵”。
一些浮游生物物种生长碳酸钙外壳(CaCO ^),改变碳的化学表面水域。当CaCO ^壳沉入深海,表面CO浓度^ 2 -减少,使方程1中的平衡反应向左边和释放二氧化碳,从而对二氧化碳产生相反的影响从生物泵。这就是所谓的生物counterpump(或碳酸盐泵)。只有10%的出口生产,然而,与CaCO ^生产,两个生物泵的总体效果是增加碳的浓度在10%相比,海洋表面深度(Murnane et al . 1999年)。
浮游生物的增长是有限的可用性,营养,和放牧。与陆生植物,高二氧化碳不能提高浮游生物的生长,因为碳已经是丰富的海洋中。中间值和深海营养丰富,但它们在大部分海洋表面耗尽,增长发生的地方。南大洋和北赤道太平洋,然而,有很高的营养但是生物生产力较低。他们被称为类食物,low-chlorophyll (HNLC)地区。他们的低生产率通常被解释为低可用性的铁(,他26章,本卷)。铁是人体必需的phy-toplankton叶绿素合成和硝酸盐还原。铁的缺乏从而限制了生产力。HNLC地区的原位实验表明,施肥的海洋铁可以增强海洋生产力。实际对出口生产的影响和潜在的持久性二氧化碳,然而,是不确定的(,他26章)。
可变性
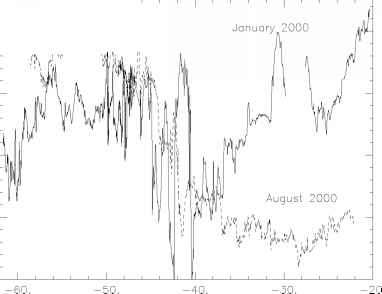
在海水中二氧化碳的分压(二氧化碳分压)和二氧化碳通量变化区域和全球海面温度(SST)时,生物生产、出口或海洋环流的变化。由于众多海洋邮轮进行自60,季节性变化如今在区域和全球尺度上建立相对较好(见综合在高桥et al . 2002年)。图12.2给出了一个例子的原位观察南印度洋在1月和2000年8月。二氧化碳分压的大的季节性变化主要是由温度控制在亚热带,但是通过生物生产和冬天混合高纬度地区(< 50°S),导致相反的在这两个地区季节性。
二氧化碳分压和海气界面二氧化碳通量的年际变化不太著名的在全球范围内。一个设变暖导致海洋二氧化碳分压增加了4%。在全球海温年际变化测量通过卫星在±0.25°C,可能导致二氧化碳通量±0.8热解色谱y-1。raybet雷竞技最新气候模型被迫通过不同的场景预测表面温度将温暖在下个世纪1.4°到5.8°C,从而减少海洋的吸收能力。
改变生物出口生产几乎直接转化为改变
380年。
360年。
340年。
320年。
300年。
380年。
360年。
340年。
320年。
300年。
纬度(程度)
图12.2。经向分布的海洋fCO2测量2000年1月(实线)和2000年8月在西南印度洋(虚线)(同时邮轮)。fCO2是理想的二氧化碳分压修正行为的二氧化碳气体。季节性fCO2幅度在南大洋,南部极地(50°S)面前,不是高达在亚热带锋区但足以从夏天二氧化碳沉到冬天二氧化碳源(Metzl et al . 2001年)。这些数据凸显了细观可变性(高点和低点1 - 10规模)在额叶区(40到45°S)主要生产的是增强在南国的夏天。
二氧化碳通量。同比变化在全球出口生产从卫星数据(图12.1)计算的±0.4热解色谱y-1。很难说将来生物出口生产将如何改变。最近的研究强调极端政权海洋生态系统的变化,要么显示(查韦斯et al . 2003年)年代际变化或趋势(Beaugrand et al . 2002年)。我们还不能估计这样的生态系统变化对二氧化碳通量的影响。我们需要理解和量化不同的浮游生物功能类型对二氧化碳的影响,估计这些类型在不同气候条件下的反应。在这一领域的研究非常活跃。卡尔et al。(1999)表明,海洋分层支持的存在浮游植物类型可以解决大气氮,但导致低出口生产。Riebe -
出售et al。(2000)表明,酸化的海洋二氧化碳浓度升高阻碍CaCO ^贝壳的形成通过浮游植物,它有两个作用:它改变碳表面化学和增加二氧化碳吸收,但降低了POC的出口能力,因为CaCO ^壳与高出口生产(克洛斯和弓箭手2002)。这些影响已经开始被纳入全球碳模型(Bopp et al . 2001;博伊德和多尼2002),但还有很长的路要走可以依靠这种模式的结果。
海洋环流的变化所带来的影响也很难估计,因为缺乏大规模的观测。我们主要依靠海洋模型来估计这一项。混合的海洋中发生的空间尺度上1米(m)到10千米(公里),一个数量级小于的分辨率当前全球碳排放模型。上升流是由风散度。从卫星风现在可以测量,但估计的不确定性和模型产生的上升流。海岸上涌在全球模型也不好解决。冰融化导致海洋分层,也会对二氧化碳通量的影响。所有的这些影响都可以估计与当前海洋模型,但这些估计的可靠性是值得商榷的。热含量的观测表明,模型能够再现观察气候趋势但不是其自然变化(莱维图斯et al . 2001;raybet雷竞技最新Bopp et al . 2002年)。
循环变化在CO2通量的影响取决于感兴趣的时间尺度。年际变化,减少中间的曝光率,从分层或减少上涌,会导致大气中的二氧化碳通量异常海洋,因为中间水域在DIC比地表水丰富。在长时间尺度,我们必须考虑到循环变化对人为二氧化碳的影响。人为的二氧化碳吸收就会减少,如果海洋分层,自然DIC的相反效果。净效应取决于大小和持续时间的变化,可以估计最可靠的模型(模型估计,看到格林布拉特和Sarmiento,第13章,本卷)。
人为二氧化碳通量在稳态的海洋
当大气中的二氧化碳增加,海洋吸收的速度,取决于大气中的二氧化碳的增加速度,而海洋混合的速度。只需要大约一年的海洋表面与大气平衡,但这需要几年几个世纪混合表面与中间值和深水海洋。因此,人为海洋中碳的储存目前达到只有~ 15%的潜在最大(图12.3)。海洋有可能吸收人为二氧化碳的70 - 85,但这需要几个世纪后排放发生(阿切尔et al . 1997年)。
大气中的二氧化碳的比例,可以与高二氧化碳被海洋吸收减少。随着越来越多的人为二氧化碳在海洋里溶解,少CO32”可以形成HCO3(方程1)。因此更多的二氧化碳保持可溶性气体的形式,它可以

图12.3。全球意思是现在的库存(实线)和潜在的最大(虚线)的人为二氧化碳的大气中的二氧化碳浓度368 ppm。两条曲线之间的差异是由于海洋的混合时间长。当前存储计算平均使用人为DIC估计世界各地。水平线表示一个标准差。观测的分析是由r . m .主要基于数据中引用Sabine et al .(1999、2002)和李et al。(2003)。
图12.3。全球平均当前的库存(实线)和潜在的最大(虚线)的人为二氧化碳的大气中的二氧化碳浓度368 ppm。两条曲线之间的差异是由于海洋的混合时间长。当前存储计算平均使用人为DIC估计世界各地。水平线表示一个标准差。观测的分析是由r . m .主要基于数据中引用Sabine et al .(1999、2002)和李et al。(2003)。
交换与大气。这种影响是巨大的,容易预测(格林布拉特和Sarmiento,第13章)。
全球海洋碳的吸收可以使用几个独立的估计方法:大气二氧化碳的测量,用813 c扩展,O2 / N2,或大气中的二氧化碳含量的空间分布;直接测量的海洋碳,表面813 c,海洋或相关;反演海洋示踪剂;和全球海洋模型。最近估计1.5和2.8之间的吸收范围包括y-1 1980 - 2000年,代表年度化石燃料排放量的四分之一到一半(表12.1)。为整个工业时期直到1994年的总吸收碳估计为111±13包括基于观测(Lee et al . 2003年),大约四分之一的人为排放雷竞技csgo总量(化石燃料+土地利用;房子et al . 2002年)。模型结果的观察。
尽管结果引用都是相互连贯的,收敛向1.9的值包括y-1 1980 - 2000年(Prentice等等。2001),中值的不确定性仍在±0.7热解色谱y-1。该方法基于大气CO2和O2 / N2,发达国家在1990年代,为减少这种不确定性提供了保证。该方法假定,
表12.1。的估计意味着海洋二氧化碳吸收包括y-1)不同 |
||
方法和最近的时间段(基于普伦蒂斯et al . 2001年,表3.4中, |
||
随着进一步的研究) |
||
时间和学习 |
方法 |
吸收 |
1970 - 1990 |
||
码头et al。(1992) |
海洋13 c库存 |
2.1±0.8 |
这个和Maier-Reimer (1996) |
海洋13 c库存 |
2.1±0.9 |
1980 - 1989 |
||
普伦蒂斯et al。(2001) |
大气中的二氧化碳和氧气/氮气 |
1.9±0.6 |
Bopp et al。(2002);Le Quere et al。(2003) |
大气CO2和O2 / N2 |
1.8±0.8 |
普拉特纳先生et al。(2002) |
大气CO2和O2 / N2 |
1.7±0.6 |
战斗et al。(1996) |
南极冰原iceb O2 / N2 |
1.8±1.0 |
奥尔et al。(2001) |
模型相互比较 |
1.85±0.35 |
麦克尼尔et al . (2003) |
海洋CFC库存 |
1.6±0.4 |
1985 - 1995 |
||
格鲁伯和基林(2001) |
海洋13 c库存 |
1.5±0.9 |
1990 - 1996 |
||
Bopp et al。(2002) |
大气CO2和O2 / N2 |
2.3±0.7 |
1994年 |
||
基于李et al。(2003) c |
海洋人为二氧化碳 |
1.9到2.4 |
库存 |
||
1995年 |
||
高桥et al。(2002) d |
海洋表面二氧化碳分压 |
2.8±1.5 |
1990 - 1999 |
||
普伦蒂斯et al。(2001) |
大气中的二氧化碳和氧气/氮气 |
1.7±0.5 |
Le Quere et al。(2003) |
大气CO2和O2 / N2 |
1.9±0.7 |
Keeling和加西亚(2002)e |
大气CO2和O2 / N2 |
1.9±0.6 |
普拉特纳先生et al。(2002) |
大气CO2和O2 / N2 |
2.4±0.6 |
Ciais et al。(1995);鞣料et al。(1989); |
大气中的二氧化碳和S13Cb |
1.8 |
Trolier et al。(1996) |
||
Keeling和派珀(2000) |
大气中的二氧化碳和S13C |
2.4 |
麦克尼尔et al . (2003) |
海洋CFC库存 |
2.0±0.4 |
海洋O2 aga通量的变化来修正 |
内容(参见文本的解释不同 |
|
估计)。 |
||
bCalculations更新普伦蒂斯et al。(2001)基于引用的引用。 |
||
cAssumes海洋和大气中的二氧化碳增加 |
遵循相似的曲线。 |
|
d纠正+ 0.6热解色谱y-1占土地 |
海洋河流流量。 |
|
e1990 - 2000。 |
||
然而,海洋氧气通量为零在过去的几年里,它就不会是这样了如果海洋变暖。必须纠正预算基于O2占海洋氧气通量。我们可以估计这个假设O2和热通量校正的比例和使用热通量估算。Keeling和加西亚(2002)估计每焦耳4.9 nanomoles的比例因子(nmol / J),从地球化学观察,而Bopp et al。(2002)和普拉特纳先生et al。(2002)估计价值高20% ~从模型的结果。海洋热含量直接估计基于数以百万计的温度资料1950 - 1996年(莱维图斯et al . 2000年)。1980年代,海洋热含量下降了0.13 X 1022 J y-1;1990 - 1996年,它增长了1.0 x 1022 J y-1。热含量测量的编译没有超出1996。
修正的碳预算1990年代很多依赖假定的热含量变化为1996 - 2000。结合温度和海平面高度观察,Le Quere et al。(2003)估计,海洋热含量增加0.55 X 1022 J y-1,平均在1990年代。相比之下,普拉特纳先生et al .(2002)认为1.24的速度X1022 J y-1观察1990 - 1995年期间保持在整个1990年代,而Keeling和加西亚(2002)使用0.6倍的速度1022 J y-1,根据模型结果包括人为强迫但没有自然变化。Bopp et al。(2002)他们只能通过1996年的预算。海洋二氧化碳水槽基于O2 / N2纠正与observational-based热含量为1.8±0.8,1.9±0.7热解色谱y-1 1980年代和1990年代,分别(Le Quere et al . 2003年)。
很难估计得多区域通量比全球吸收的二氧化碳(彩色版9从Gloor et al . 2003年)。大气中的二氧化碳测量东西梯度小,转化为区域通量估算大气反演中巨大的不确定性。海洋二氧化碳分压的测量有很大的不确定性与缺乏相关的测量相对于区域和颞可变性,估计通量,我们无法准确量化气体交换。海洋反演避免由于气体交换,但不确定性与不确定性估计海洋运输和数据字段。海洋反演估算区域二氧化碳通量测量的DIC, O2,碱度,营养。全球海洋模型普遍认同彼此但是显示不同地区吸收了100%的平均通量(奥尔et al . 2001年)。尽管这些不确定性,所有方法显示出出气的二氧化碳的吸收在热带和中高纬度地区(彩色版9),符合我们的理解底层的过程(图12.1)。各种估计之间最大的分歧是在南部海洋,观察在哪里稀疏的(彩色版9)。
不确定性和未来的研究需要
自第一次报告政府间气候变化专门委员会在1990年,科学家们信心的海洋吸收~ 2包括y-1大大增加了。raybet雷竞技最新初步估计,基于模型的结果,现在还支持广泛的观察。这个估算的准确性,然而,很难约束。估计基于大气测量受阻需要准确地量化过程忽视了到目前为止(即。、土地变化813 c分馏、海洋O2 / N2出气);估计的基础上海洋测量的大时空采样频率要求;和海洋反演和模型,需要准确地量化垂直运输和海洋中混合过程。另一方面,可以在很大程度上减少不确定性区域通量估计和在全球的趋势。更好的限制区域海洋通量可以大大改善大气反演。南大洋worst-constrained地区。46个°S南部的海洋覆盖了16%的世界海洋和通风大约一半的深海水域。区域的测量表明,该地区的水槽1包括y-1(高桥et al . 2002),但大气反演看到水槽只有0.4热解色谱y-1(格尼et al . 2002年)。两种方法之间的差异可能由于缺乏本地测量在冬季(Metzl等等。1999年,2001年;图12.2),但这还有待在整个盆地的规模。未来的南大洋吸收二氧化碳的反应是最大的不确定性来源之一的预测未来大气中的二氧化碳(Friedlingstein,第十章,这卷)。
这里大部分的不确定性引用仅适用于平均状态。1980年代和1990年代之间的区别似乎是更健壮的均值估计要么十多年(Le Quere et al . 2003年)。监测站(热、蝙蝠)(韦恩et al . 1999;贝茨et al . 2002年)和重复调查(井上et al . 1995;费利et al . 2002年)测量二氧化碳或者DIC的变化在过去的几十年中由于高度仪器精度和DIC标准(Dickson 2001)。理解和预测这些趋势,重要的是要理清流程驱动的变化,特别是海洋环流和混合的趋势,这已经被证明能够影响DIC(贝茨et al . 2002年)。为此我们需要大规模监测海洋环流的变化对海洋生物地球化学和混合及其影响。启动漂移ARGO浮标在海洋将提供杰出的连续测量温度和盐度的海洋。附件这些漂浮的O2传感器将提供极有价值的信息对海洋生物地球化学的变化。
科学家们对海洋生物之间的相互作用的理解,biogeochem-istry,和物理需要改进。这个不足是最好的说明了这样一个事实,我们不能解释是什么导致了80 ppm (ppm)的变化在结冰期的大气中的二氧化碳,虽然它是肯定的,原因在于海洋bio-geochemistry(乔斯和学徒,第七章,这卷)。这个问题必须解决有效的海洋生物地球化学模型。然而现在的模型很少考虑生态系统动力学和不同的浮游生物功能类型的角色。差距在专家的垂直混合的理解阻碍整个生物地球化学循环的发展模式。检查观察年际年代际变化的表面碳循环和中间也是一个重要的任务检测和了解目前趋势或转移和验证模型能够再现这些变化。这次考试可以实现只有在长期时间序列,表面进行测量,维护和盆地规模部分动力学和在不同的海洋生物地球化学领域,如果数据交付和合成的观察发生在国际层面上。直到显示模型可以复制glacial-interglacial大气中的二氧化碳生物地球化学和现代趋势的变化,我们的信心在CO2通量的预测将是有限的。
确认
我们感谢j·格林布拉特提供表12.1;n格鲁伯,c . Sabine j . l . Sarmiento和两个匿名评论家建设性意见;SeaWiFS项目工作人员(NASA代码970.2)提供的数据图12.1;船上工作人员r . V Marion-Dufresne提供数据图12.2(同时项目支持的法国国家科学研究所de l 'Univers,皮埃尔西蒙拉普拉斯,研究所,研究所Polaire Paul-Emile维克多);r . m .关键人为编制的全球估计DIC图12.3;和m . Gloor提供彩色版9。
文献引用
阿切尔D。h . Kheshgi,大肠Maier-Reimer。1997年。多个时间尺度的化石燃料二氧化碳中和。《地球物理研究快报》24 (4):405 - 408。
贝茨:R。,a·c·Pequignet r·j·约翰逊,和n格鲁伯》2002。短期大气二氧化碳在亚热带模式水下沉的北大西洋。自然420:489 - 493。
战斗中,M。m·本德,t .苗圃,P P黝黑色,j·h·巴特勒,j . w . Elkins j·t·埃利斯t·康威n, P . Lang和公元克拉克》1996。大气中的气体浓度在过去一个世纪以空气从积雪在南极。自然283:231 -235。
战斗中,M。m·本德P P黝黑色,j·w·c·白,j·t·埃利斯t·康威和r . j . Francey》2000。全球碳汇及其变化,从大气O2和S13C推断。科学287:2467 - 2470。
Beaugrand、G、P c·里德,f . Ibanez说j·a·林德利和m·爱德华兹,2002。重组的北大西洋海洋桡足动物生物多样性和气候。raybet雷竞技最新科学296:1692 - 1694。
Behrenfeld, m . J。,1997 P·g·法尔。。消费指南浮游植物初级生产力模型。湖沼学和海洋学42:1479 - 1491。
Bopp、L。,p . Monfray o . Aumont J.-L。杜福瑞斯h . LeTreut g . Madec l . Terray和j·奥尔》2001。气候变化对海洋出口生产的潜在影响。raybet雷竞技最新全球生物地球化学循环15:81 - 99。
Bopp、L。c . Le Quere m•海曼著,a·c·曼宁,p . Monfray。2002。raybet雷竞技最新海洋气候导致氧气通量:对当代碳预算的影响。全球生物地球化学循环16日gb001445 doi: 10.1029/2001。
博伊德,P W。,s·c·多尼。2002。造型区域海洋深海生态系统对全球气候变化的响应。raybet雷竞技最新《地球物理研究快报》29日gl0l4l30 doi: 10.1029/2001。
查韦斯,f . 1 !j·r·瑞恩,e . Lluch-Cota和m . Niquen》2003。从凤尾鱼、沙丁鱼和背部:太平洋数十年的变化。科学2999:217 - 221。
Ciais, P。P黝黑色,j . w .白色,m . Trolier r . Francey j .贝瑞·d·兰德尔,P卖家,j . Collatz和d .丢弃。1995。分区的海洋和陆地吸收的二氧化碳所推断的S13C测量从NOAA全球气候监测和诊断实验室空气采样网络。raybet雷竞技最新地球物理研究100:50515070杂志》上。
迪克森a . 2001。海洋测量的参考资料。海洋学14 (4):21 - 22。
费利,R。,j . Boutin c . Cosca y Dandonneau, j . Etcheto h .井上,M。Ishii, c . Le Quere d·麦基m . McPhaden n . Metzl a .泊松和r . Wanninkhov。2002。季节性和二氧化碳在赤道太平洋的年际变化。深海研究49:2443 - 2469。
Gloor, M。:格鲁伯,j·l·Sarmiento c . l . Sabine r·费利,和c Redenbeck》2003。现在和工业化前的初始估计海气CO2通量模式基于海洋内部碳测量和模型。《地球物理研究快报》30 (1),gl015594 doi: 10.1029/2002。
格鲁伯,N。,2001 c, d·基林。。改进估计二氧化碳同位素海气界面不均衡的:对人为的海洋吸收二氧化碳的影响。《地球物理研究快报28:555 - 558。
格尼,k . R。,r . m .法律,丹宁,p . j .雷纳·d·贝克,p . Bousquet l . Bruhwiler中州。陈,P Ciais,风扇,即y Fung) m . Gloor m .这个k . Higuchi j .约翰t Maki s Maksyutov k . Masarie P . Peylin m·普莱瑟公元前Pak, j·安德森,j . Sarmiento s .田口t .高桥,C.-W。元2002。对健壮的地区估计二氧化碳的源和汇使用大气传输模型。自然415:626 - 630。
这个,M。和大肠Maier-Reimer。1996年。之间的关系及其碳同位素海洋吸收二氧化碳。全球生物地球化学循环10:89 - 110。
房子,i . J。,即c·普伦蒂斯和c Le Quere》2002。最大的影响未来的重新造林或砍伐森林对大气中的二氧化碳。全球变化生物学8:1-6。
井上,h . Y。,h . Matsueda h . Ishii k .故事m .大臣浅沼的,和y Takasugi》1995。长期趋势的二氧化碳的分压(二氧化碳分压)在西方北太平洋表层海水,1984 - 1993。忒勒斯47 b: 391 - 413。
卡尔,d . M。r·勒特里尔,l .领袖j·多尔,j . Chrisitan和d﹒荷伯》1999。固氮的作用在亚热带北太平洋海洋生物地球化学循环。自然388:533 - 538。
基林,r F。和h·e·加西亚。2002。海洋O2库存的变化与最近的全球变暖。自然科学院99:78487853学报》上。
基林,c, D。,s . c . Piper。2000。年际变化的大气中的二氧化碳和13 CO2交流海洋与陆地生物圈和从1978年到2000年。三世。模拟源和汇。斯克里普斯海洋研究所的参考00-14。圣地亚哥:加州大学斯克里普斯海洋研究所的。
克洛斯,C。,2002 d·阿切尔。。协会沉没有机质与各种类型的深海矿物镇流器:对雨比率的影响。全球生物地球化学
周期,gb0011765 doi: 10.1029/2001。
法律、大肠。P·g·法尔w·o·史密斯Jr . h . Ducklow和j·j·麦卡锡。2000年。在公海对出口温度的影响生产。全球生物地球化学循环14:1231 - 1246。
李,K。南达科他州。崔G.-H。公园,r . Wanninkhof郭宏源。彭,r . m .钥匙,c . l . Sabine r·a·费利,j·l·Bullister和f . j . Millero》2003。一个更新的人为二氧化碳库存在大西洋。全球生物地球化学循环(提交)。
Le Quere C。o . Aumont l . Bopp, p . Bousquet p . Ciais r . Francey m .这个c·d·基林r·f·基林h . Kheshgi p . Peylin s c . Piper i c·普伦蒂斯,p . j .雷纳。2003。二十年的海洋二氧化碳水槽和可变性。忒勒斯55:649 - 656。
莱维图斯。j . i安东诺夫t·p·波伊尔,和c·斯蒂芬斯。2000。海洋变暖的世界。科学287:2225 - 2229。
莱维图斯。t . l . j . i安东诺夫j . Wang Delworth, k·w·迪克森和a . j .西兰花。2001。人为气候变暖的地球的气候系统。raybet雷竞技最新科学292:267 - 270。
麦克尼尔,我。r . j . Matear r m键,j·l·Bullister和j·l·Sarmiento》2003。基于全球人为二氧化碳被海洋吸收含氯氟烃数据集。自然299:235 - 239。
Metzl, N。a、b . Tilbrook和泊松》1999。年度fCO2周期和海气界面二氧化碳通量的子任务南冰洋。忒勒斯51 b (4): 849 - 861。
Metzl, N。c .深色,a . Jabaud-Jan答:泊松,Schauer b。2001。夏季和冬季海气CO2fluxes在南大洋。第688 - 685页。二氧化碳第六届国际研讨会论文集,卷2。日本筑波:全球环境研究中心。
Murnane, R。,和c, j·l·Sarmiento Quere》1999。海气界面二氧化碳通量的空间分布和碳的两半球间的运输海洋。全球生物地球化学循环13:287 - 305。
奥尔,j . C。,e . Maier-Reimer Mikolajewicz, p . Monfray j·l·Sarmiento j . r . Toggweiler n . k .泰勒,j·帕尔默,n .格鲁伯c . l . Sabine c . Le Quere r m键,j . Boutin》2001。人为碳吸收量的估计从四个三维全球海洋模型。全球生物地球化学循环15:43-60。
普拉特纳先生,G.-K。f·乔斯,t . f .储料器》2002。全球碳预算的修正由于氧海气通量的变化。全球生物地球化学循环,gb001746 doi: 10.1029/2001。
普伦蒂斯,i C。g·d·法夸尔,m . j . r . Fasham m . l . Goulden m .这个和v . j . Jaramillo h . s . Kheshgi c . Le Quere r . j .斯科尔斯和d·w·r·华莱士。2001。碳循环和大气中的二氧化碳。2001年第237 - 183raybet雷竞技最新页。在气候变化:科学依据(工作组的贡献我的政府间气候变化专门委员会的第三份评估报告),由j·t·霍顿编辑y叮,d . j . Griggs m . Noguer p . j .范德林登戴x, k . Maskell和c·a·约翰逊。剑桥:剑桥大学出版社。
码头,p D。b . Tilbrook, c . s . Wong》1992。海洋化石燃料二氧化碳的吸收:Car-bon-13证据。科学256:74 - 79。
Riebesell U。桑德凡,b·罗斯特P Tortell, r .齐伯,f·莫雷尔。2000。海洋浮游生物减少钙化的反应增加了大气中的二氧化碳。自然407 (6802):364 - 367。2
Sabine, C。,L。R . m .关键,k·m·约翰逊f . j . Millero j·l·Sarmiento d . w . R。
华莱士和c·d·韦恩。1999。人为二氧化碳的印度洋库存。GlobalBiogeochemical周期13:179 - 198。
Sabine, c . L。r·a·费利,r . m .钥匙,j·l·Bullister f . j . Millero郭宏源k . Lee。彭,b . Tilbrook t .小野,和黄》2002。人为二氧化碳的分布在太平洋。全球生物地球化学循环,gb001639 doi: 10.1029/2001。
高桥,T。s c·萨瑟兰,c·斯威尼泊松,n . Metzl b . Tilbrook n .贝茨r . Wanninkhof r·a·费利,c . Sabine j . Olafsson和y Nojiri》2002。全球海洋空气CO2flux基于气候海洋表面二氧化碳分压,和季节性生物和温度的影响。深海研究II 49:1601 - 1622。
黝黑色,P P。、t·j·康威和t . Nakazawa》1989。纬向分布的源和汇的大气二氧化碳来自表面观察和大气传输模型。地球物理研究杂志94:5151 - 5172。
Trolier, M。,j·w·c·怀特,P P .黝黑色,k . a . Masarie a和P拥有德国。1996。监测大气CO2-measurements从NOAA的同位素组成全球空气取样网络。地球物理研究杂志101:25897 - 25916。
Wanninkhof, r S。,多尼、t .高桥和w·麦吉利。2001年。使用平均风的影响在全球海气二氧化碳通量。351 -356页。在水表面气体传输,由m·a . Donelan编辑w . m .庄士贤e·s·萨尔兹曼和r . Wanninkhof。AGU的地球物理专题系列卷》127。华盛顿特区:AGU的出版社。
韦恩,c, D。,中州。李、f·t·麦肯齐和d·m·卡尔。1999。不断上升的海洋表面溶解无机碳在夏威夷海洋时间序列的网站。海洋化学60:33-47。
这篇文章有用吗?