安德鲁·G喷泉W贝瑞里昂
介绍
气候变化的观点在更新世和raybet雷竞技最新全新世仅仅十年前非常不同。冰芯记录的收集和详细分析从格陵兰岛和南极洲早在1990年代中期,分别在这时期气候变化的集体观点已经发生了巨大的变化。raybet雷竞技最新更新世期间,b.p至少早在450000年。,突然和剧烈的温度波动是经常出现,而非特例(Mayewski et al . 1996, 1998;小et al . 1999年)。更新世期间,这些快速和大的气候波动,最初确定的冰芯记录,已在海洋和验证湖泊沉积物(债券et al . 1993;格林et al . 1993),表明大规模重组(全球)半球气候在很短的时间(Mayewski et al . 1997年)。raybet雷竞技最新类似的气候波动,但较小的振幅,也发生在全新世(O ' brien et al . 1995;债券et al . 1997;Arz et al . 2001年)。是什么生物反应温度的变化、降水、大气化学?我们必须回答这个问题,如果我们要理解的世纪——millennial-scale影响气候对生态系统的结构和功能。raybet雷竞技最新
由于极地地区被认为是全球气候变化的放大器,这些地区非常适合调查生态系统的反应,被认为是可能的在温带地区,小规模的气候变化。raybet雷竞技最新我们对过去气候变化在南极洲来自不同类型的代理记录,包括冰芯、地质和海洋(里昂et al . 1997年)。然而,很明显,南极洲沿海可能回应海洋、大气,
南极东部冰盖,
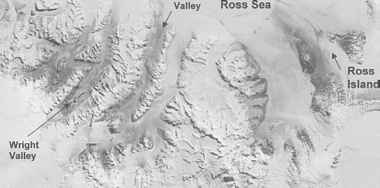
图16.1的陆地卫星图像麦克默多干谷。南极东部冰盖在左下角的照片和罗斯海冰的右上角照片被雪覆盖。深色阴影识别裸露的土壤干旱的山谷。
冰sheet-based气候“司机”,因此无冰的区域,如麦克默多干谷,可以应对气候变化比之前认为的更复杂的方式(r波,unpubl。raybet雷竞技最新2001年数据)。自启动麦克默多干谷长期生态研究计划(MCM)在1993年,有浓厚的兴趣不仅在今天的动态的生态系统,而且在通过过去的气候变化对生态系统产生的遗产。在这一章里,我们检查当前的干谷生态系统结构和功能的角度来看我们的工作集中在泰勒谷。从这种理解我们研究生态系统应对气候变化的变化在过去的27000年,强调过去的气候条件的重要性,对当前生态系统功能。
网站描述
麦克默多干谷的长期生态研究(MCM)站点位于南极大陆的边缘(77.5°S, 163°E)在更大的区域称为南部维多利亚地(图16.1)。只有2%的南极冰是免费的(首席伊朗研究员Drewery et al . 1982年)和麦克默多干谷是最大的无冰地区面积约2000平方公里的无雪的大陆(Chinn 1988)。山谷欠他们的存在横贯山脉那块冰流从南极东部冰盖(图16.1)。几叶的冰盖穿透山脉和山谷。此外,许多小高山杯子-

图16.2西方泰勒谷看西方。邦尼湖在左下角。注意无冰缘(护城河)在低端的湖。中央在后台是休斯冰河反曲刀山。极左是Sollas冰川,冰川的右边的影子是火山灰锥,是用Wilch et al .(1993)到目前为止冰川活动。(由托马斯Nylen照片)
《形式在山上和流到谷底。山谷的特点是rocky-sandy土壤缺乏血管植被,从而产生鲜明的景观(图16.2)。常年冰雪覆盖的湖泊,这是美联储从短暂的小溪原始冰川融水存在于几乎所有的山谷(喷泉et al . 1999年)。黑暗的极地气候地区经raybet雷竞技最新验不断在冬至和持续的阳光在仲夏。气温平均约-17°C,冬季最小约-40°C和夏季最大几度零上(克洛et al . 1988年)。
降水发生如雪,可以在任何时候。每年降雪的值约10厘米,水当量(键1980),其中大部分是输给了升华(Chinn 1993)。干谷展览非常低的生物学生物多样性和短食物链(Priscu et al . 1999;维吉尼亚和墙1999)。土壤社区是有限的几门,包括轮虫,缓步类,nema-todes、原生动物、真菌和细菌(Freckman和弗吉尼亚州1998)。线虫是最大的食肉动物的山谷。蓝藻的流支持社区,真核藻类,苔藓(麦克奈特et al . 1999年)。湖泊主机只有微生物和底栖微生物垫(沃顿商学院等。1993年,Priscu et al . 1999年)。小微生物群居住在自然(冰穴)冰川融化洞(沃顿商学院et al . 1985年)和湖里的冰(Priscu et al . 1998年)。虽然
-

- 图16.3地图泰勒谷

生活在南极洲的高纬度地区,存在干谷主机最后功能的陆地生态系统之一,溪流,湖泊和土壤物理和生物互联。
MCM主要集中在泰勒谷,东至west-trending谷长约34公里,宽12公里(图16.3)。东,泰勒谷麦克默多海峡,开放和西部被泰勒冰川,从南极东部冰盖流(图16.1)。海拔从海平面到2000在山区,北部和南部边界的山谷。谷底的海拔范围从海平面到约60米。MCM于1993年发起的,研究编译中可以找到Priscu在生物科学(1998)和(1999)。理解气候变化如何影响泰勒谷生态系统,一个更完整的物理描述山谷,生态系统,和他们的交互是必需的。
物理环境
从结构上看,干谷地区作为一个刚性块三级初以来,5000万年前(菲茨杰拉德et al . 1986年)。大地构造隆起在泰勒谷(Wilch et al . 1993年),这是重要的生态过程的山谷,已经过去257万年相对较慢。在上新世,小火山锥爆发泰勒谷中间的中间区域,显然产生很少或没有灰存款。因为锥是坐落在山谷两侧,一般海拔在300米以上,并远离谷底,他们在谷生态的角色可能是小,这里不考虑。干谷隆起受到密切关注,因为可能影响区域气候变化(贝伦特和库珀1991)。raybet雷竞技最新最近考试Wilch et al。(1993)认为,根据其结构特点、火山锥不可能爆发。因此,视锥细胞的年龄和高程提供限制的值的提升。如果隆起发生在过去的260万年里,它不可能超过300。
将干谷划分为沿海地貌模型,中间,和内陆地区(军火商和丹顿1996)。这些分歧大致相符,至少自中新世时期(Wilch et al . 1993),低,中间,和高海拔。位于沿海地区表现出现代土壤运动(如泥流,冰楔多边形的模式),深度ice-cemented沉积物小于50厘米。的地形和土壤都小于12000岁,因为他们是由late-Wisconsin形成冰川沉积物(解决)。Midelevation中间区域表现出相对较低的土壤水分。冰川融水是罕见的在这个区域,因此河流是少见期间除了罕见的极端的热空气的温度。Water-induced斜坡运动(泥流)仅限于一个潜在水源的地区如冰川附近或每年形成雪补丁。中间区域的景观不是动态的,气候条件有利于保护沙漠人行道和杆多边形。基于钾氩约会原位灰存款,中部斜坡运动一直最小泰勒谷在过去710万年(1996年-马尔尚和丹顿)。海拔较高的内部区域,发现800米以上,基本上没有土壤水分,融水完全缺席。没有活跃层冻土因为土壤缺乏冰。过时的火山灰存款,被困和埋在土壤中的热收缩裂缝或保存在沙漠上,表现出年龄在10 - 15几百万年(军火商和丹顿1996)。火山灰层的沉积构造表示没有斜坡运动或接触水以来沉积的时间。 Taylor Valley is completely encompassed by zones 1 and 2 because of its relative proximity to the coast compared to other dry valleys. Nonetheless, the age of the landscape surfaces is quite old compared to its temperate counterparts: It ranges in age from 12,000 years at the valley bottom to 7.1 million years on the upper valley walls.
更新世以来,重要的表面改性的低海拔(区域1)泰勒谷,像所有的干谷,已经由冰川活动,包括水从冰川径流。河谷低地区域1的土壤,由冰川形成存款或事件直接导致冰川活动。的比率风成修改似乎很小。军火商和丹顿(1996)日期近地表灰尘沉积在风高10 - 15数百万年海拔区,这表明沙漠人行道大大抑制明显风蚀或沉积。区域1的土壤含有海洋和湖泊有机物质和中级水平较低,和石内的在高海拔地区主导有机质来源(伯京et al . 2000年)。
建立的历史背景下理解所有风景是至关重要的生态系统(Swanson et al . 1988),但在MCM尤其如此
在过去气候变化真正决定生态系统现状。由于极地位置,MCM的主要干扰生态系统气候,和景观格局主要由气候、生物,流程。
raybet雷竞技最新
被认为是一个干旱的山谷极地沙漠。在很大程度上南极洲,降水非常低。测量在赖特山谷,毗邻泰勒谷,表明每年降雪范围从0.6到10厘米水当量(布罗姆利1985)。降水是最大的最近的海岸和减少内陆(键1980;喷泉et al . 1999年)。基于雪深测量在地面上的泰勒谷经过一个夏天暴风雪和积雪深度测量在泰勒谷冰川在海拔200 - 300米(喷泉et al . 1999年),沉淀积累的梯度下降-0.06厘米km-1(水当量),距离海岸。通常,雪融化之前升华物,使水文小贡献。只会在这些地方雪积累重要的深度和不受风,如洼地,流渠道或冰川的利润率,它做出贡献,如果瞬态,水文的山谷。
在泰勒谷,温度范围从几度零上年末12月或1月冬天到-45°C(克洛et al . 1988年,多兰et al . 2002 b)。年平均气温在硅谷范围从-16°C到-20°C。控制温度的重要因素之一下吹的风南极东部冰盖流。这些foehn-type绝热地温暖的风从高海拔和流动可以提高空气温度在15 - 20分钟10度。因此,冬季气温也在一定程度上控制的频率下降风(Doran et al . 2002 b)。除了气候变暖,这些风大大降低冰表面的湿度和显著提高升华(克洛et al . 1988年)。除了重力的影响,山谷通常刮风。月平均风速范围从2到4 m s - 1在泰勒谷(克洛et al . 1988年)。
冰川
几乎所有的干谷相对较小的冰川高山冰川,几平方公里的区域。在泰勒谷,冰川来自北边的仙宫范围和反曲刀山在南边(图16.1),积雪超过损失的升华和风力侵蚀。泰勒冰川是最大的冰川谷,定义其西部边界,从南极东部冰盖流。大约三分之一的泰勒谷脊分裂的定义山脉是冰覆盖(喷泉et al . 1998年)。的冰川正在极地冰川与衬底基地冻结。更大的冰川终止在垂直冰崖约20米高。与冰温带冰川,冰是非常干净和自由的碎片。因为冰是远低于冰点,冰川融水仅限于表面和冰川的边缘。这些特征与温带高山冰川,在他们的熔点和不冻结他们的床。相反,他们回避底物。冰川融水的温带进入人体,内部流动,最后从冰川下的隧道中冰川边缘(喷泉和困境1998)。
这些极地冰川的质量交换相对较小,而温带冰川。我们的观察表明,10 - 30厘米的雪积累上区和6到15厘米的丢失消融区。这些值是一致的结果从先前的研究在邻赖特(牛和Carnein 1970;Chinn 1980)。雪的上游冰川寒冷和干燥,和没有融雪直接观察到。质量损失在雪地上区是升华,因为没有发生融化。在低海拔冰川(ice-exposed)区,质量损失是由升华和融化。的结果加拿大冰川表明,在夏季,升华占40 - 80%的冰质量损失,其余是输给了融化(Lewis et al . 1998年)。自1993年以来,融水生产是有限的下边缘在几百米的冰川冰边缘。当然,没有观察到厚的冰川海拔250 - 500米以上。
水文
冰川融水流的形式流底部的冰崖。这些流是短暂的,通常每年流十周(麦克奈特et al . 1999年)。他们运输水、沉积物和营养终端湖泊(图16.4)。如前所述,降雪在谷底不大大加剧流,因为它通常升华物融化之前(Chinn 1980)。然而,冬天的雪堆积在山巅的冰川末端或流渠道导致水流在早春消失之前,初夏(喷泉et al . 1998年)。信道和流流连续多年冻土,发生在浅深几十厘米(麦克奈特et al . 1999年)。因此,地下水流动可能是局限于近地表hy-porheic区,饱和区毗邻和河道。因为浅深度和缺乏横向地下水流入,hyporheic区可以扩展横向两侧数米的通道。
提要的冰川融水流通常溶质含量很低,1 - 10的微摩尔(|)/化学物种(里昂et al . 1998年)。冰川的溶质含量控制,化学部分的积雪,但另有由回收溶质吹到谷底的冰川表面(里昂等人在出版社)。冰川融水化学空间模式的初步分析表明,它是由沉积物的模式在冰川(m·特兰特unpubl。数据,2001)。溪水的质量流量增加一到两个数量级之前到达湖的解散和盐化学风化作用过程内的通道和hyporheic区(里昂et al . 1998;Gooseff 2002)。蒸发损失河道也有助于增加溶质的浓度(b·沃恩unpubl。数据,1993)。
三个主要湖泊占领泰勒山谷的底部(弗里克塞尔湖,霍尔
-
 into the lakes (black). McMurdo Sound is the ocean area.")
- 图16.4地图泰勒谷中溪流,排水的冰川湖泊(黑)(灰色)。麦克默多海峡是海洋区域。
 into the lakes (black). McMurdo Sound is the ocean area.")
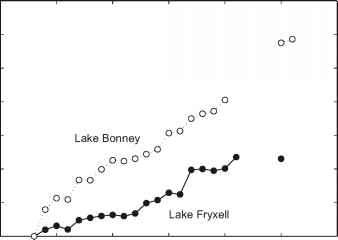
邦尼)除了无数的小湖泊和池塘点的景观。所有的湖泊和池塘有永久性冰盖(厚3 - 6米),和较小的水体可能完全冰冻的冬天。如前所述,这些湖泊是终端,失去水只有通过升华的冰表面蒸发从狭窄的边缘开放水域的四周每个湖在夏末。与其他终端湖泊(例如,大盐湖),这些湖泊在进水量小的变化很敏感。自1970年代开始的测量记录由Chinn(1993),一般干谷中所有的湖泊已经上升。直到最近,湖泊放缓或停止上升(图16.5)。湖霍尔,水平下降在过去的几年中,以应对夏季凉爽的天气和夏天降雪增加(喷泉et al . 1999;多兰et al . 2002年)。因为常年冰有效地覆盖了湖水,没有风能产生的电流(霍斯1983)。湖水之间的气体交换和气氛因此限制(沃顿商学院et al . 1986)光穿透减少到湖水(Howard-Williams et al . 1998年)。泰勒谷的主要湖泊因为盐度梯度密度分层(腾出空间和Priscu 1998)。一些混合发生在夏季湖边缘,冰融化和局部湍流可以传播到湖(米勒和艾肯1996)。湖水的运动只限于慢水平运动。融化湖边缘还允许有限与大气交换气体。此外,水流通过融化边缘进入湖泊,提供营养和水分的湖(泰勒et al . 1998年)。
作为OL
1970年
1975年
1980年
1985年
1990年
1995 2000
图16.5湖泰勒谷水平变化。
1975年
1980年
1985年
1990年
1995 2000
图16.5湖泰勒谷水平变化。
土壤
卵石砾(1到20厘米直径)路面表面覆盖粗糙,但变量,一点一点凝聚力和土壤质地和有机质含量(Bockheim 1997;坎贝尔et al . 1998年)。冻土无处不在的山谷,在土壤通常是发现在10 - 30-cm-thick活跃层。在泰勒谷,土壤质地为主95 - 99%的粗粒度砂(伯京et al . 2000年)。土壤质地高可变性的结果冰川作用在硅谷,随后进行了描述。土壤的有机质含量不超过0.03%的有机碳含量(伯京et al . 2000年)。土壤的水分含量较低,随距离海岸。在沿海地区,土壤水分平均重量约1%;更远的内陆山谷地板上土壤水分降至0.5%(坎贝尔et al . 1998年)。规模较小的模式土壤水分取决于靠近水源,例如湖泊和短暂的流。此外,冬天雪积累,通常单薄而且很不协调,提供一个暂时的水源。然而,这些来源的影响不超过10米(坎贝尔et al . 1998;麦克奈特et al . 1999年),和95%的土壤得不到液态水(坎贝尔et al . 1998年)。这些土壤水分取决于短暂的积累的雪,融化之前通常升华,或升华ice-cemented冻土深度(坎贝尔et al . 1998; McKay et al. 1998).
生态系统
生态系统的干谷是相对孤立的,因为高carolina pagli山脉的山谷和内部水分开山谷。之间缺乏运输山谷的线粒体DNA分析表明了土壤线虫,Scottnema lindsayae。据说这种线虫的基因会相当统一,因为它是由风和流很容易分散,但土壤样本从五个不同的山谷mito-chondrial方面表现出显著差异,表明种群之间的基因流动受到限制(Courtright et al . 2000年)。在山谷内,土壤之间的联系、流和湖泊生态系统在温带地区相比相对较弱的山谷。在泰勒谷,坡面流土壤和地下水是不存在的,因此,连接到只有通过hyporheic区河流或湖泊河流每年几周期间的相互作用。
一个整合的过程,虽然弱,是风。风成过程运输沉积物和有机物质穿过山谷。风乘火车生物体(1999年弗吉尼亚和墙)从土壤和侵蚀藻垫和苔藓流渠道(麦克奈特et al . 1999年)。在湖泊底栖藻垫从湖底的浮力引起的累积气体垫(帕克et al . 1982),成为冻湖冰的表面。通过持续的冻结在底部和顶部的升华,这些垫通过冰和运输成为暴露在地表风力侵蚀的地方。所有这些材料都是穿越山谷和沉积在土壤、河道和湖泊。通常发现一些藻丛英联邦冰川在海拔300米。我们假设净泥沙运输和有机物质向海岸由于高速风谷事件引起的重力风流动的冰原。尽管down-valley梯度增加有机碳(1999年弗吉尼亚和墙),观察到局部修改,作为解释为气候事件(我们在本章后面讨论),也会导致这一趋势。
干谷的极地气候包括一raybet雷竞技最新个黑暗的漫长的冬天,非常寒冷,没有地表水。在这段时间内生态系统之间的联系都断绝了。当然风继续分散有机质,但生态系统不再是集成和唯一功能的生态系统,湖泊,完全隔离。每个系统采用的策略为了熬过冬天,在隔离。因此,在这样一个环境中生物之间的联系系统一定会很脆弱,因为他们是切断了季节。土壤ne-matodes输入状态低湿休眠,它允许他们生存的冬天没有水和低于冰点的温度(1971克)。他们已经知道生存在这种状态下60多年(Freckman 1986)。流苔藓和藻垫也成为冻干后停止供应融水,以及流停止流动。再湿润之后,一些垫在10 - 20分钟开始光合作用(Vincent和Howard-Williams 1986;霍斯et al . 1992年)。最近的实验表明,藻垫没有经验的水流了25年的重新激活一个完整的微生物生态系统在一两周内达到1.5公里流再湿润(麦克奈特et al . 1999年)。冬天,湖泊成为独立于外部环境开放水域的边缘内的湖泊和常年湖冰冻结。剩下的冬天,湖环境是完全与外界隔绝任何交换。此外,一些存在于极地秋季和春季,而完全黑暗发生在冬天。Phy -
toplankton湖泊,这取决于光进行光合作用,替代能源。研究南极湖泊,尤其是在干燥山谷,表明一些浮游植物物种将细菌摄取,有机碳的来源,在低光照条件下(罗伯茨和Laybourn-Parry 1999;Laybourn-Parry et al . 2000年)。这种混合营养的策略允许浮游植物忍受冬天的黑暗。我们对冬至的理解生物过程的湖泊是不完整的,因为物流的限制,到目前为止,杜绝在冬天的研究。在春季浮游植物生长完全由触发增加太阳辐射,因为垂直混合基本上是一片空白。生物量和浮游植物生产一般通过1月下旬提高整个夏天。
世纪——Millennial-Scale气候变化raybet雷竞技最新
raybet雷竞技最新气候变化的干谷已基本推断从过去的冰川地貌证据的位置(例如,Stuiver et al . 1981;丹顿et al . 1989年)和湖水平高度(例如,Stuiver et al . 1981;大厅和丹顿1995,2000)。此外,概要文件的化学浓度的湖泊被用来推断过去湖经费(威尔逊1964;里昂et al . 1998年)。最近,从冰核同位素结果(史泰格et al . 2000年)和温度测量在基岩水井(g .克洛unpubl。数据,1993年冰川)和(克洛和沃丁顿1996)被用来更直接推断出该地区的气候变化。raybet雷竞技最新我们总结millennial-scale气候变化基于这些证据,使用冰raybet雷竞技最新芯数据作为我们的基准年表。
冰核从泰勒穹顶,从泰勒谷约140公里。尽管取得了一些核心从大陆(例如,Jouzel et al . 1987;摩根et al . 1997;Mayewski et al . 1996;罗格朗和Mayewski 1997),这个特殊的核心是最接近干谷,提供更好的信息远比核心区域气候的变化。事实上,结果从泰勒圆顶核心与核心结果Vostok和伯德(Blunier et al . 1997, 1998),它位于两端的南极大陆远离泰勒圆顶。气候数据推断出从其他核心从格陵兰岛的阶段,而泰勒圆顶的数据核心阶段。这些差异归因于空间变量深海洋流运输热南大洋和低纬度地区之间(史泰格et al . 1998年)。
各种地球化学分析泰勒圆顶核心已经完成,但我们把精力集中在这些结果的重要性的气候干谷所总结的史泰格et al . (2000)。raybet雷竞技最新图16.6显示了氧同位素的推断气温和积雪从冰芯记录。开始大约6万(可以)前(日历年)开始缓慢冷却,使过去冰川最大。在同一时期,积雪减少,达到一个最低在上次冰河最大值。快速变暖,被称为“B0lling-Aller0d事件,发生大约15可以前终止的冰河时期。随后,新仙女木期冷却
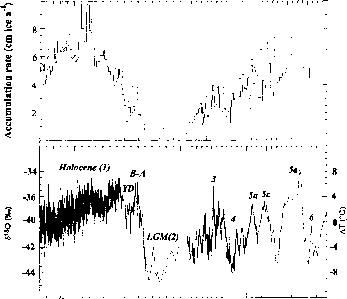
图16.6:堆积速率在cm中每年的冰(cm冰a - 1)在泰勒圆顶基于10数据。底部:氧同位素值(S180)泰勒穹顶(左轴)和推断温度变化(在)(右轴)。是几千年的时间尺度现在之前。注意,在20000年时间尺度的变化。这些数据从史泰格等。2000年,分别为图7和图6。
图16.6:堆积速率在cm中每年的冰(cm冰a - 1)在泰勒圆顶基于10数据。底部:氧同位素值(S180)泰勒穹顶(左轴)和推断温度变化(在)(右轴)。时间表在几千年之前。注意,在20000年时间尺度的变化。这些数据从史泰格等。2000年,分别为图7和图6。
事件之前气温下降4°C左右回到温暖B0lling-Aller0d值约11可以前。大量冷却事件可以发生在9.5和6.5可以前。从6.5的B0lling-Aller0d事件通过冷却事件可以前,通常积累增加。一般的降温趋势和减少积累一直占据着晚全新世。晚全新世温度趋势的细节(几个可以)不可以在泰勒圆顶冰核,因为这个时候潜在nontemperature影响同位素数据(沃丁顿珀耳斯通讯,2000)。幸运的是,地下地球温度测量在泰勒谷,钻孔和温度随深度变化的基础上,获得了表面温度的记录(1998年克洛)。一般冷却保存在硅谷从4可以前约1可以。从那时起,空气温度加热。
这些气候趋势造成地区冰盖的变化范围和湖的水平。去年冰川最大的冷却,南极冰盖开始扩大和推进。约23.8 (C14)可以前(~ 27可以)罗斯冰架从海洋进入了泰勒谷(大厅和丹顿2000;Stuiver et al . 1981年)。冰架封锁了外海的山谷,使一个内陆湖,称为沃什伯恩湖,深约200米。湖中存在直到大约8.3 (C14)可以前(~ 9.5可以)当山谷的冰架消退和湖水域排水的山谷。硅谷大约4可以以来冷却1.2°C;这可能导致了湖泊的主要减少干谷,达到其最大下降0.9 - -1.2可以前(威尔逊1964;里昂et al . 1998年)。湖霍尔完全干燥或不存在~ 2之前可以前(里昂et al . 1998年)。从那时起,湖泊也加过气温上升2°C在过去的1000年里。这些最近的山谷的变化与其他推断该地区的气候变化。raybet雷竞技最新 For example, the "penguin optimum," associated with rookery abundance and a warmer climate, occurred between 3 and 4 kyrs ago and terminated about 3 kyrs ago (Baroni and Orombelli 1994). The rookeries were reoccupied starting about 1200 years ago.
人类在1903年第一次参观了干谷当罗伯特·f·斯科特的西部之旅包括泰勒冰川湖邦尼路缘带下来。政党的成员测量的宽度最窄一点的湖,后来被Chinn(1993)来估计一个湖的水平。自1903年以来的湖泊已经上升,通常表明气候变暖。raybet雷竞技最新这一趋势仍在继续从1970年代,当Chinn(1993)开始测量,通过1980年代。变薄的湖冰建议2°C变暖在1980年代,尽管其他因素可能是重要的(麦凯et al . 1985年)。这种变暖可能导致融化,径流增加,进而导致增加湖的水平。从1986年到2000年,气温下降了0.7°C每十年,从冰川径流已经放缓,湖水平普遍下降停止上升,在某些情况下(多兰et al . 2002;韦尔奇et al .,这本书第十章)。
生态系统响应
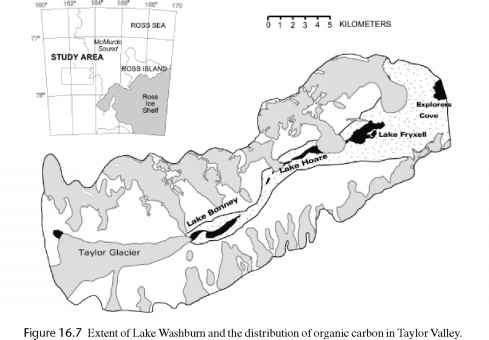
沃什伯恩湖的形成大约27可以前形成了一座湖,从山谷口在西端泰勒冰川(图16.7)。Phyto-plankton水柱和底栖藻垫显然居住这扩大湖因为伯京et al。(2000)发现的湖泊和海洋模式的土壤中有机碳来源横断面与前湖。沃什伯恩湖的湖总生物量在时间比目前的力度要大得多。这个生物的有机碳在土壤仍然存在由伯京表示的工作。因此,当前在谷中碳排放速度非常慢。迄今为止,它认为wind-transported有机碳(湖泊微生物垫,石内的社区,和基于流的垫子和苔藓)在硅谷是一个碳转移的主要来源。尽管实时观测表明,这是真的,这个过程是有争议的意义的相关性lacustrine-derived有机碳和湖的程度沃什伯恩(伯京et al . 2000年)。由于缺乏更高的植物和动物,土壤生物显然是持续的有机碳土壤留下的沃什伯恩湖。因此,干谷湖泊连接到土壤在时间而不是在空间。
一旦罗斯冰架从泰勒谷约9.5可以前,冰不再封锁了山谷,和沃什伯恩湖干涸,留下小湖泊
面对低斑点沿着谷底。气候冷却raybet雷竞技最新4可以前开始和冰川的河流速度低于要求维持湖泊。正因为如此,大小湖泊开始减少。他们通过~ 1000年前达到最小。如果湖霍尔存在之前,它已经完全消失,残留的盐被吹走(里昂et al . 1998年)。我们假定溪流生态系统在很大程度上没有和土壤生态系统已经大大减少了。是否所有陆地生物群进入了一个长期低湿休眠无法确定。弗里克塞尔湖和邦尼蒸发卤水池,浓缩盐和营养等有机和无机碳。从那时起,气候温暖和更大的融水湖通量增加。raybet雷竞技最新新鲜的融水没有混合浓缩卤水池,由于密度的差异,和浮在上面,捕获的盐水深度。 Diffusion fluxes from the concentrated brines at depth have been used to obtain the refilling age of ~1 kyr bp (Wilson 1964; Lyons et al. 1998).
邦尼湖的浮游植物的研究表明,其生产力和生物量增加太阳辐射穿透冰层覆盖在赛季早期(Priscu et al . 1999年)。这首先发生在增加浅水区冰的表面下。总的来说,只有不到0.1%的溶解碳池在营养生成区是由水流(Priscu et al . 1999年)。随着光照强度的增加与南国夏至的方法,主要地区的生产力变化从表面15米的水柱底部营养生成区(图16.8)。这种转变的结果足够的光线到达
-
 Photosyntheti-cally available radiation. (b) Primary productivity, (c) Chlorophyll a. (From Priscu et al. 1999)")
- 图16.8水东叶邦尼湖的列属性。(一)Photosyntheti-cally可用辐射。(b)初级生产力,(c)叶绿素a。(从Priscu et al . 1999年)
 Photosyntheti-cally available radiation. (b) Primary productivity, (c) Chlorophyll a. (From Priscu et al. 1999)")
相对的高通量的深度无机和有机碳,通过分子扩散下面,浓缩池的营养(Priscu 1995;Priscu et al . 1999年)。我们认为,浮游植物继续生产在这个深度到南极秋天后流停止流动。生产力的深度可能会改变更浅水平再次响应低光照水平的南极的冬天。不幸的是,我们缺乏测量在此期间和冬季由于后勤方面的约束。在任何情况下,很明显,当前湖泊生态系统强烈受制于过去的气候事件。大约1000年前撤军的湖泊集中湖泊营养物质进入池现代浮游植物现在所依赖。
过去的痕迹在当前生态系统功能的影响被称为遗产(沃格特et al . 1997年)。在我们的例子中,遗产涉及气候的变化及其对湖泊的影响大小。raybet雷竞技最新以前大湖低地土壤提供养分,残余的当前湖泊paleolakes供给营养。至关重要的气候干谷是由于其遗产极端的环境、生物多样性低,短的食物链。极端极性环境减少可用的水和能源生态系统,大大减缓了养分循环的速度。低生物多样性和短食物链使生态系统直接依赖于物理环境,这样一些缓冲区存在和轻微的气候变化的生态系统的响应是立竿见影。这是说明了当前湖泊初级生产力下降由于气候冷却(Doran et al . 2002年)。raybet雷竞技最新降温趋势降低了冰川meltwa-ter产生的大小,进而降低了河流和湖泊营养物质通量。湖泊水位已停止上升,在某些情况下,正在下降。应该强调,冰芯和井眼温度数据显示,这些巨大的变化在湖的大小由年度变化2°C或更少。我们再次强调,在温带设置这些微小的变化,但在极地环境中这些相对较小的差异放大带来非常大的环境响应。泰勒谷这是由于,在某种程度上,这个生态系统是如此紧密依赖的变化状态的水,在南国的夏天,很小的温度变化可以极大地影响水文系统。
一般生态系统问题和总结
在研讨会上世纪Millennial-Scale气候变化和生态系统响应,作为长期生态研究计划的一部raybet雷竞技最新分的所有科学家会议雪鸟,犹他州(2 - 4 2000年8月),几个将军生态系统被问到的问题。这些都是在这里总结麦克默多干谷lte。
先前存在的条件影响气候事件的意义吗?我们的食物链是由微生物群应对微环境变化,变化明显在短距离和随着时间的推移。缺乏高等植物群落的领带我们陆地生态系统更紧密地地质结构比温带地区。贫瘠的环境空间联系较弱和减缓地貌变化随着时间的推移提高连杆(遗留)(摩尔黑德et al . 1999年)获得者来到卡罗莱纳州。
这种影响是直接或级联的吗?我们目前的理解是,影响是直接的,因为短的食物链。可怜的联系,否则缓冲区没有直接影响。
是生态系统响应之前完成下一个事件?不同大小的事件发生在不同的时间尺度,每个都有自己的响应时间。然而,当前遗留的沃什伯恩湖事件(27000年前)是与当前遗留的湖的撤军1200年前。极慢骑自行车的营养和地貌变化的速度,我们怀疑生态系统响应相互叠覆上,不是之前完成下一个事件发生。
系统返回它吗初始状态之后的事件吗?我们不知道,但答案取决于我们目前在摄动静态平衡状态,即存在一个恒定状态的系统可以返回,还是系统处于一种动态平衡的状态,没有恒定状态,系统不同常数的值。考虑到相对较短的直接观察记录,完整记录过去的事件,和缓慢的速率变化的山谷,告诉这个时候是不可能的。然而,我们可以说,当前系统摄动,鉴于最近披露的重要遗产。
结论
麦克默多干谷的生态系统表现出强烈和明确的依赖过去的气候条件。相对较大的气候变化有集中营养物质不能在“正常”或静态条件下获得raybet雷竞技最新。由于营养物,能源有限公司环境的干谷,过去浓度的营养发挥重要作用在当前生态系统结构和功能。这个发现把人的注意力吸引到了需要理解过去的气候条件,至少在极端极性环境,解释当前的生态系统。干谷中的生态系统当然是联系薄弱,土壤和湖泊出现行为完全独立。只有通过我们对遗留我们意识到系统的理解在一个非常现实的时间和方式至关重要。
此外,我们观察一个极性放大,小生态系统响应气候变化产生很大的变化。raybet雷竞技最新能量限制在干谷一直潜在的水供应被锁在冰冻的水库周围的冰川。小的变化在夏季冰川的表面能量平衡,导致表面温度高于熔点上升,形成一个较大的水通量。相反,一个小变化,导致降低温度低于熔点显著减少水生产。干谷的夏季气温徘徊在熔点,这高度非线性响应温度变化是一个山谷的正常状态。虽然我们检查过去的气候对生态系统响应的影响不明确解决这一过程中,我们知道从我们这个领域的经验,这种非线性响应是所有观察到的变化的核心。沃什伯恩湖的创建必须发生在局部能量允许水平冰的温度温暖和维持融化。相反,能级在湖撤军必须充分下降减少冰温度低于冰点的夏天。
这些研究成果有什么提供理解生态系统的能量,富含水分环境吗?遗产,包括气候、人为和火山活动,可能是重要的空间和时间尺度。每个遗留的意义取决于的大小变化和景观和生态系统的响应时间,以适应这一变化,并返回整个系统回到先前存在的条件。也许通过了解自然变化的影响和停留时间,我们可以预测人为改变的影响。此外,我们的研究可能有助于帧生活如何生存的极端事件如bollide影响或病情“雪球”地球。当生态联系都被切断,一些弹性是由遗留影响储存营养,否则不可用。
确认本章将不可能没有我们的洞察力和劳作lte同事和研究生。这一章是献给他们。我们还欣赏南极的精湛的努力支持Associates提供的后勤支持,让这个项目成为可能。该项目资金是由美国国家科学基金会提供,极地办公室项目,格兰特麦克默多干谷:A寒冷的沙漠生态系统,OPP 9211773, OPP 9813061;和自然遗产的作用在极地沙漠生态系统结构和功能:OPP麦克默多干谷lte项目9810219,OPP 0096250。
引用
Arz, h·W。美国姑娘,j . Patzold Rohl。2001。Millennial-scale表层和深层水流的变化在西部热带大西洋与北半球高纬度地区的气候raybet雷竞技最新在全新世期间。地质29:239 - 242。
巴罗尼C。,G. Orombelli. 1994. Abandoned penguin rookeries as Holocene paleocli-matic indicators in Antarctica. Geology 22:23 -26.
贝伦特,j . C。,A. Cooper. 1991. Evidence of rapid Cenozoic uplift of the shoulder escarpment of the Cenozoic West Antarctic rift system and a speculation on possible climatic forcing. Geology 19:315-319.
生物科学。1999年。麦克默多干谷(专题)。BiosScience 49:9591019。
Blunier, T。,9 others, 1997. Timing of the Antarctic cold reversal and atmospheric CO2 increase with respect to the新仙女木事件事件。《地球物理研究快报24:2683 - 2686。
Blunier, T。,10 others, 1998. Asynchrony of Antarctic and Greenland climate change during the last glacial period. Nature 394:739-743.
键,G。,13 others, 1993. Evidence for massive discharges of icebergs into the North Atlantic Ocean during the last glacial period. Nature 360:245-249.
Bockheim, j·g . 1997。从南极寒冷的沙漠土壤属性和分类。美国土壤科学学会杂志61:224 - 231。
键,G。w .淋浴,m . Cheseby r . Lotti p . Almasi p . de Menocal p . Priore h·卡伦,i Hajdas, g . Bonani。1997。一个普遍的在北大西洋全新世和冰川气候millennial-scale周期。raybet雷竞技最新科学278:1257 - 1266。
布罗姆利,a . a . 1985。天气观测:赖特山谷,南极洲。信息发布11。新西兰的气象服务。惠灵顿,新西兰。
牛,C。,C. R. Carnein. 1970. The mass balance of a cold glacier: Meserve Glacier, south Victoria Land, Antarctica. International Association of Hydrological Sciences 86:429446.
坎贝尔,i B。g·g·c·克拉里奇,d。坎贝尔和m . r .阻止。1998。土壤环境的麦克默多干谷,南极洲。在Priscu, j . c(编辑)在沙漠极地生态系统动力学。美国地球物理联盟。南极研究系列2:297 - 322。
伯京,m . B。弗吉尼亚州,r . a . c·p·张伯伦·d·h·墙。2000。土壤有机质的来源在泰勒谷,南极洲。生态81:2377 - 2391。Chinn, t . j . 1980。维多利亚冰川平衡干谷地区,土地,南极洲。国际水文科学协会126:237 - 247。Chinn, t . j . 1988。维多利亚“干谷”的土地,在r·s·威廉姆斯和j·g·Ferrigno。南极冰川的世界地图,卫星图像。 U.S. Geological Survey Professional Paper 1386-B:B39-B41. Chinn T. J. 1993. Physical hydrology of the Dry Valley lakes. Pages 1-52 in W. J. Green and E. I. Friedmann, editors. Physical and Biological Processes in Antarctic lakes. Antarctic Research Series 59. American Geophysical Union, Washington, DC. Clow, G. D. 1998. Reconstructing past climatic changes in Greenland and Antarctica from borehole temperature measurements [abs]. EOS—Transactions of美国地球物理联盟79:F833。小丑,g D。c·p·麦凯,g·m·西蒙斯Jr .)和r·a·沃顿,Jr .) 1988。气候观测和预测升华利率湖霍尔,南极洲。气候学杂志1:715 - 728。
小丑,g D。,E. D. Waddington. 1996. Acquisition of borehole temperature measurements from Taylor Dome and dry valleys for paleoclimate reconstruction. Antarctic Journal of the United States 31:71-72. Courtright, E. M., D. H. Wall, R. A. Virginia, L. M. Frisse, J. T. Vida, and W. K. Thomas. 2000. Nuclear and Mitochondrial DNA Sequence Diversity in the Antarctic Nematode Scottnema lindsayae. Journal of Nematology 32:143-153. Crowe, J. H., 1971. Anhydrobiosis: An unsolved problem. American Naturalist 105:563571.
丹顿,g . H。j·g . Bockheim s . c·威尔逊和m . Stuiver。1989。威斯康辛州和全新世冰川历史,内心的罗斯湾,南极洲。第四纪研究31:151 - 182。
多兰,p . T。j . c . Priscu w·b·里昂,j·e·沃尔什a·g·喷泉d·m·麦克奈特·d·k .运算r . a .弗吉尼亚·d·h·墙,g·d·克洛h . g . Fritsen c·p·麦凯a·p·帕森斯,2002。南极气候raybet雷竞技最新冷却和陆地生态系统响应。自然415:517 - 520。
多兰,p . T。c·p·麦凯,g·d·克洛·g·l·达纳·a·g·喷泉,t . Nylen w·b·里昂,2002 b。raybet雷竞技最新麦克默多干谷气候观测,南极洲,1986 - 2000。地球物理Research-Atmospheres杂志》上。107:1-12。首席伊朗研究员Drewery d J。、s·r·乔丹和大肠养家糊口。1982。南极冰盖的测量属性:表面配置,冰层厚度、体积和基岩特征。年报的冰川学2:834 - 891。菲茨杰拉德,p·G。, M. Sandiford, P. J. Barrett, and A. J. W. Gleadow. 1986. Asymmetric extension associated with uplift in the Transantarctic Mountains and Ross Embayment. Earth and Planetary Science Letters 81:67-78. Fountain, A. G, G. L. Dana, K. J. Lewis, B. L. Vaughn, and D. M. McKnight. 1998. Glaciers of the McMurdo Dry Valleys, Southern Victoria Land, Antarctica. Pages 65-76 in J.
c . Priscu编辑器。在极地沙漠生态系统动力学:南极洲麦克默多干谷。南极研究系列72,美国地球物理联盟,华盛顿,
喷泉,a G。,12 others. 1999. Physical controls on the Taylor Valley ecosystem, Antarctica. BioScience 49:961-971.
喷泉,a G。,J. S. Walder. 1998. Water flow through temperate glaciers. Reviews of Geophysics 36:299-328.
Freckman, d . 1986。在土壤生物的生态脱水。页157 - 168 a . c . Deopold编辑器。膜、新陈代谢和干燥的有机体。纽约伊萨卡康奈尔大学出版社,。
Freckman D。,R. Virginia. 1998. Soil biodiversity and community structure in the McMurdo Dry Valleys, Antarctica. Pages 323-336 in J. C. Priscu, editor. Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica. Antarctic Research Series 72, American Geophysical Union, Washington, D.C.
Gooseff m . N。w·b·里昂,a·e·布卢姆和d·m·麦克奈特。2002年。风化反应和hyporheic交换控制流在冰川融水水化学流在麦克默多干谷。水资源研究,38:15-1-15-17。
格林,。g·l·雅各布森,Jr .) w·a·瓦b . c·s·汉森和k . a . Maasch。1993。50000年的记录从佛罗里达气候振荡和raybet雷竞技最新与海因里希的时间相关的事件。科学261:198 - 200。
大厅,B。,G. H. Denton. 1995. Late Quaternary lake levels in the Dry Valleys, Antarctica. Antarctic Journal of the United States 30:52-53.
大厅,B。,G. H. Denton. 2000. Radiocarbon chronology of Ross Sea Drift, eastern Taylor Valley, Antarctica: Evidence for a grounded ice sheet in the Ross Sea at the last glacial maximum. Geografiska Annaler 82A:305-336.
霍斯,i . 1983。湍流及其后果两个冰雪覆盖的南极湖泊浮游植物的发展。英国南极调查公报60:69 - 81。
霍斯,我。、c . Howard-Williams和w·f·文森特。1992。干燥和恢复蓝藻的南极的垫子。极地生物学12:587 - 594。
Howard-Williams C。a·施瓦兹,即霍斯,j . c . Priscu。1998。光学性质的湖泊的麦克默多干谷。页189 - 204 j . c . Priscu编辑器。在极地沙漠生态系统动力学:南极洲麦克默多干谷。南极研究系列72,美国地球物理联盟,华盛顿特区
Jouzel, J。c . Lorius j . r .小c . Genthon n . i Barkov v . m . Kotlyakov和v . m .彼得罗夫》1987。Vostok冰芯:连续同位素温度记录在过去的气候周期(160000年)。自然329:403 - 407。
钥匙,j . r . 1980。气温、风、降水和大气湿度在麦克默多地区。地质系出版17号(南极数据系列号9),新西兰惠灵顿维多利亚大学的。
Laybourn-Parry, J。、e·c·罗伯茨和e·m·贝尔》2000。Mixotrophy在南极湖泊浮游原生动物之间的生存策略。页面33-40 w·戴维森,c . Howard-Williams和p . Broady编辑器。南极生态系统:为更广泛的生态模型的理解。卡克斯顿出版社,克赖斯特彻奇,新西兰。
罗格朗,M。,P. A. Mayewski. 1997. Glaciochemistry of polar ice cores: A review. Reviews of Geophysics 35:219-243.
刘易斯,k . J。,a·g·喷泉和g·l·达纳。1998。能量平衡研究加拿大冰川,泰勒谷,麦克默多干谷,南极洲。年报的冰川学27:603609。
里昂,w . B。,p . a . Mayewski l . r . Bartek和p·t·多兰。1997。raybet雷竞技最新气候的历史麦克默多干谷自最后一个冰河最大值。第15 - 22页在c . HowardWilliams w·b·里昂和霍斯,编辑器。在南极生态系统过程IceFree风景。Balkema,鹿特丹,荷兰。
里昂,w . B。s w·泰勒,r·a·沃顿Jr .) d·m·麦克奈特和b·h·沃恩。1998。全新世晚期干燥霍尔和弗里克塞尔湖,湖南极洲麦克默多干谷。南极科学10:247 - 256。
里昂,w . B。k·a·韦尔奇,a·g·喷泉g·d·达纳·b·v·沃恩·d·m·麦克奈特,在出版社,表面glaciochemistry泰勒谷南部维多利亚地,南极洲,流化学及其关系。水文过程。
里昂,w . B。k·a·韦尔奇,c . a . Nezat d·m·麦克奈特k·克里克,j . k . Toxey j . a . Mastrine》1998。在弗里克塞尔湖流域化学风化速率和反应,泰勒谷:比较温和的江河流域。页147 - 154 c . Howard-Williams w·b·里昂和霍斯,编辑器。在南极生态系统过程无冰的风景。Balkema,鹿特丹,荷兰。
Marchant, d R。,G. H. Denton. 1996. Miocene and Pliocene paleoclimate of the Dry Valley region, southern Victoria Land: A geomorphological approach. Marine Mi-cropaleontology 27:253-271.
Mayewski, p。,13 others. 1996. Climate change during the last deglaciation in Antarctica. Science 272:1636-1638.
Mayewski,公共广播l·d·米克尔,m . s . Twickler s甲沟炎,问:杨,w·b·里昂,1998 m·普伦蒂斯。。北半球高纬度地区大气环流的主要特性和强迫使用长110000年glaciochemical系列。地球物理研究杂志102:26345 - 26366。
麦凯,c·P。g·克洛,r·a·沃顿Jr .)和s·w·斯奎尔斯。1985。常年冰冻的湖面上的冰的厚度。自然313:561 - 562。
麦凯,c·P。m·t·梅隆,依弗里德曼,1998。土壤温度稳定ice-cemented地面麦克默多干谷,南极洲。南极科学,10:31-38。
麦克奈特,d . M。d . k .他a s阿尔及尔,a . Bomblies p . a . Conovitz和c·m·泰特。1999。干谷流在南极洲:等待水生态系统。生物科学49:985 - 995。
米勒,l·G。,G. R. Aiken. 1996. Effects of glacial meltwater inflows and moat freezing on mixing in an ice-covered Antarctic lake as interpreted from stable isotope and tritium distributions. Limnology and Oceanography 41:966-976.
Moorehead, d . L。p·t·多兰,a·g·喷泉w·b·里昂·d·m·麦克奈特j . c . Priscu r . a .弗吉尼亚州,1999 d·h·墙。。生态遗产:麦克默多干谷的对生态系统的影响。生物科学49:1009 - 1019。
摩根,我诉。t . d . c . w . Wolley j . Li研究,1997 w·斯金纳,m·f·菲茨帕特里克。。网站信息和初步结果深钻在法律穹顶,南极洲。杂志的冰川学48:3-10。
O ' brien, s R。,p . a . Mayewski l·d·米克·d·a·米斯m . s . Twickler, s . i甲沟炎。1995。的复杂性全新世气候raybet雷竞技最新从格陵兰冰芯和重建。科学270:1962 - 1964。
帕克,b . C。,通用西蒙斯,Jr . g . f .爱,r·a·沃顿Jr .)和k . g . Seaburg》1982。去除有机和无机物从南极湖泊蓝藻垫的空中逃脱。《藻类学18:72 - 78。
小,j . R。,18 others. 1999. Climate and atmospheric history of the past 420,000 years from the Vostok ice record, Antarctica. Science 286:2141-2144.
Priscu, j . c . 1995。湖泊的浮游植物养分缺乏麦克默多干谷,南极洲。淡水生物学34:215 - 227。
Priscu, j . c(编辑)。1998年。在沙漠极地生态系统动力学,麦克默多干谷,南极洲。南极研究系列72。美国地球物理联盟,华盛顿特区
Priscu, j . C。,c . h . Fritsen e·e·亚当斯,s . i Giovannoni h·w·Paerl, c·p·麦凯p·t·多兰d·a·戈登,调查局Lanoil和j·l·平克尼。1998。常年南极湖泊冰:生活在极地沙漠绿洲。科学280:2095 - 2098。
Priscu, j . C。c·f·沃尔夫·c·d·塔卡克斯c . h . Fritsen j . Laybourn-Parry e·c·罗伯茨解决,和w·b·里昂。1999。常年冰雪覆盖的南极湖泊碳转换。生物科学49:997 - 1008。
罗伯茨,e . C。,J. Laybourn-Parry. 1999. Mixotrophic cryptophytes and their predators in the Dry Valley lakes of Antarctica. Freshwater Biology 41:737-746.
腾出空间,r . H。,J. C. Priscu. 1998. Physical limnology of the McMurdo Dry Valleys lakes. Pages 153-187 in J. C. Priscu, editor, Ecosystem dynamics in a polar desert: The McMurdo Dry Valleys, Antarctica. Antarctic Research Series 72, American Geophysical Union, Washington, D.C.
史泰格,e . J。e·j·布鲁克,j·w·c·白,c . m . Sucher m·l·本德美国j .雷曼e . d .沃丁顿d·l·莫尔斯,1998 g·d·克洛。同步在南极气候变化和北大raybet雷竞技最新西洋。科学282:92 - 95。
史泰格,e . J。d·l·莫尔斯·d·沃丁顿m . Stuiver p . m . groot p . a . Mayewski m . s . Twickler, s . i甲沟炎。2000。威斯康星阶泰勒和全新世气候历史的冰芯穹顶,西方罗斯raybet雷竞技最新湾,南极洲。Geografiska记录82:213 - 235。
Stuiver M。g·h·丹顿,t·j·休斯和j·l·Fastook》1981。海洋的历史在南极洲西部冰原在过去冰川作用:一个工作假说。页319 - 436 g·h·丹顿和t·h·休斯,编辑。最后一个巨大的冰盖。Wiley-Interscience,纽约。
Swanson, f·J。t . k . Kratz:凯恩,r . g . Woodmansee》1988。地形对生态系统的影响模式和流程。生物科学38:92 - 98。
泰勒,s W。p·g·库克,a z对接,j·m·托马斯·p·t·多兰和w·b·里昂。1998。的证据深循环在两个常年冰雪覆盖的南极湖泊。湖沼学和海洋学43:625 - 635。
文森特,w·F。,C. Howard-Williams. 1986. Antarctic stream ecosystems: Physiological ecology of a blue-green algal epilithon. Freshwater Biology 16:219-233.
弗吉尼亚,r。,D. H. Wall. 1999. How soils structure communities in the南极干谷。生物科学49:973 - 983。
沃格特,e . a . 1997。生态系统:平衡科学与管理。斯普林格出版社,纽约。
沃顿商学院,r。,c·p·麦凯g·d·克洛和d·t·安徒生》1993。常年冰雪覆盖,对南极湖泊生态系统的影响。页53 - 70 w·j·格林,依弗里德曼,编辑。物理和生物过程在南极湖泊。南极研究系列59岁的美国地球物理联盟,华盛顿特区
沃顿商学院,r。公元前,c·p·麦凯帕克、小g·m·西蒙斯。1986。氧气的预算一个常年冰雪覆盖的南极干谷湖。湖沼学和海洋学31:437 - 443。
沃顿商学院,r。,c·p·麦凯g·m·西蒙斯Jr .)和帕克。公元前1985年。冰川冰穴。生物科学35:499 - 503。
Wilch, t·R。g·h·丹顿,d . r .勒克斯,1993 w·c·麦金托什。。有限Pliocent冰川范围和表面隆起中泰勒谷,南极洲。Geografiska记录75:331 - 351。
威尔逊,a . t . 1964。气候变化的证据从化学扩散麦克默多干谷1200年前。自然201:176 - 177。
继续阅读:总结气候变化和生态系统响应的Jornada lte的网站
这篇文章有用吗?
读者的问题
-
Donata4个月前
- 回复