模型的概述的地步
模拟使用一个基本的模型可能会从一组初始条件指定作物种植,土壤中的水分和养分的初始状态,和所需的参数代表作物的生理特点。
-

- 图4.1为一个假设的地步模型流程图,日常时间步

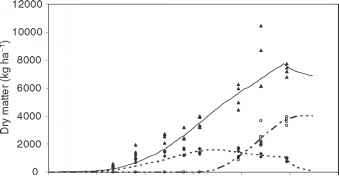
为一年生作物,模型遍历一系列子程序,估计植物或土壤处理每小时或每天的基础上,输出中间值在指定的时间间隔(图4.1)。在每一个周期中,作物模型检查是否已达到成熟或丰收的数据,在这种情况下,产量和各种各样的汇总数据可以输出。通常情况下,这些模型的输出由一系列连续的曲线代表不同的植物或环境变量随时间变化的。图4.2给出了这样的例子的输出种植系统模型(CSM) -CROPGRO模型的单一的农作物菜豆种植在哥伦比亚卡利市。
数学上,模型集成了一个微分方程组描述利率随时间变化。作物的预测(状态)变量可能包括器官的干质量、叶面积、根长度和土壤中的垂直分布,发展进步,土壤水分和土壤养分浓度的个体层或视野。在实践中,解析解公式过于复杂,所以它们是集成数值用几秒钟的时间步骤非常详细的模型或每小时,每天,当发现在大多数模型。
数以百计的地步模型创建了。其中许多是由一个科学家或小团队为单个研究的目的。大多数这些模型现在只能发现在文献中,虽然他们的算法可能会持续在更新模型。没有正式的系统命名法,在某些情况下,一个由不同群体独立模式已经被修改,导致混乱版本指出版物。表4.1列出了四个家庭的模型,在气候变化研究中广泛使用。raybet雷竞技最新
基本流程代表在这里描述的地步模型主要是参照一个假想的平均。大多数模型实际上报告输出在土地面积的基础上,对应于一个社区的植物相同的“平均”。一些模型可以模拟基因混合物,或同一物种的不同物种,包括杂草。
上衣
iffim |
||
V——«f。——* 1 |
■■■蒸腾■——土壤蒸发——潜在的外星人
■■■蒸腾■——土壤蒸发——潜在的外星人
50 200 150 100 0
50 200 150 100 0

——厂房吸收
图4.2仿真在帕尔米拉,投资增长的菜豆哥伦比亚使用CSM-CROPGRO-Drybean模型。(a)顶部的干物质变化(总地上材料)、树叶和谷物。从个人复制符号表明观测数据。(b)每日最大和最小温度。(c)蒸腾、土壤蒸发从和潜在蒸散。(d)氮被植物通过浸出或丢失
20 40 60
几天后种植
——厂房吸收
图4.2仿真在帕尔米拉,投资增长的菜豆哥伦比亚使用CSM-CROPGRO-Drybean模型。(a)顶部的干物质变化(总地上材料)、树叶和谷物。从个人复制符号表明观测数据。(b)每日最大和最小温度。(c)蒸腾、土壤蒸发从和潜在蒸散。(d)氮被植物通过浸出或丢失
表4.1的地步模型用于气候变化研究的例子raybet雷竞技最新
的名字
作物物种
描述和引用
示例应用程序
CropSyst
CSM-CERES
CSM-CROPGRO
史诗
大麦、玉米、高粱、大豆、小麦等
大麦、玉米、谷子、高粱和小麦
常见的豆、蚕豆、花生、大豆等豆类、棉花等
玉米、小米、大米、高粱、大豆、小麦等
辐射利用效率。每天的时间步。允许种植序列(Stockle et al . 2003年)
辐射利用效率。每天的时间步。允许种植序列(琼斯et al . 2003;Hoogenboom et al . 2004年)
Farquhar-type光合作用与hedge-row模型计算每一个光拦截。认为经济增长和维护呼吸。允许种植序列(琼斯et al . 2003;Hoogenboom et al . 2004年)辐射利用效率。每天的时间步。允许种植序列,可以耕作和土壤侵蚀模型的影响(威廉姆斯et al . 1989年)
旱作小麦澳大利亚东南部
班巴拉族花生,花生,玉米,高粱和大豆在喀麦隆
(Tingem et al . 2008年)
美国玉米和冬小麦
州(Alexandrov和Hoogenboom 2000)
小麦和玉米在伊比利亚半岛
(Minguez et al . 2007年)
玉米生产在非洲和拉丁美洲
美国(琼斯和桑顿2003)
美国大豆和花生
(Alexandrov和Hoogenboom 2000)
大豆在奥地利东北部
(Alexandrov et al . 2002年)
大豆在魁北克南部,加拿大
(臂章和辛格,2008)
在美国中西部玉米、大豆和小麦,考虑不同的空间尺度上raybet雷竞技最新气候和土壤(东方国家的人et al . 2001年)玉米、高粱、小米、大米和木薯在尼日利亚(Adejuwon 2006)
4.2.1开发准备
发展包括植物使用的流程安排等重要增长变化的出苗率,花的形成,晶粒生长迅速,发病或晶粒生长,通常被认为是生理成熟。这种生活历史可以被解释为一系列的阶段划分的阶段,所以建模方法通常被称为“发展阶段”(里奇和NeSmith 1991)。每个阶段的特征是一个持续时间表达的生理时间,这是数学上类似于热时间,成长度天或热量单位但可能包括光周期的影响,春化或其他流程。时间代表了植物所需的最短时间进步在最佳条件下从一个阶段到另一个。每天(或者每小时),工厂认为进展时间(Dt),发展速度估计的潜在增长率(DP)和潜在的利率调整等因素对温度(T)、光周期(P)和水赤字(W):
率调整因素通常从0到1为了发展低于最高速度缓慢,但强调诸如水赤字可以用来加速开发,导致因素超过一个值为1。另一种方法,在建模阶段发展,尤其常见谷类是使用叶数量为主要指标的发展进步。虽然术语不同,潜在的生理相似(Jamieson et al . 2007)。
物候学是如何建模的细节有很大的不同与作物的生物物种和决策模型的开发人员有关如何表示特定的反应。对于温度,决定涉及的选择曲线的温度变量和规范描述给定的假设形状的生理反应。常见的假设是,每个作物有“温度”,低于它并不生长或发展和一个“最适温度”,允许的最大速度增长或发展。温度可能会观察到或从每小时值估计,每日平均,或平均或加权调整各种礼仪。当然,作物不应对每天的最大或最小值温度;它暴露在温度(和所有其他环境条件)在连续的基础上。模型使用简化的温度数据和相关方程。土壤温度通常是用来控制萌发,幼苗出现,谷物,早期发展以来拍摄的国王仍然接近土壤表面。
响应曲线变化从简单的“破棍子”模型非线性函数如β函数。红衣主教在这些反应温度确定过渡点。除了前面描述的基础和最优的温度,模型不同supra-optimal温度的影响是如何表示的。最简单的方法是假定最大发育速率持续高于指定的最佳。交替,可能认为一个致命的温度下降,考虑发展的最高温度,或最大
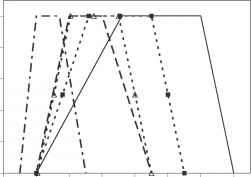
-Pre-anthesis开发。
——叶增长
————光合作用
——春化
图4.3温度响应函数的例子假定CSM-Cropsim-CERES小麦模型。pre-anthesis发展曲线是叶生长、光合作用、春化,晶粒生长。所有的反应都是基于每天的平均温度
-Pre-anthesis开发。
——叶增长
————光合作用
——春化
图4.3温度响应函数的例子假定CSM-Cropsim-CERES小麦模型。pre-anthesis发展曲线是叶生长、光合作用、春化,晶粒生长。所有的反应都是基于每日平均温度速率可能持续到第二个最佳上面率降低致死温度(图4.3)。这个领域需要进一步研究,特别是当它涉及到预计的温度上升。
非专业人员,不同方法建模发展似乎是不科学的。差异的原因是复杂的,反映基本困难一般模拟植物对天气状况的响应。最重要的是,虽然空气或土壤温度可以测量准确,植物在一个社区的经历一个复杂、波动温度环境。温度传感对于一个给定的过程可能驻留在一个特定的组织,如顶端分生组织的春化(演唱和Amasino 2004)。模型常常假设空气温度报告最近的气象站接近地上作物平均温度,但往往大树冠内温度梯度(德斯贾丁斯et al . 1978年)。另一个挑战是,温度响应包括昼夜节律(内部生物钟)的植物,从研究和结果在恒定的温度下或从简单的常量日夜政权可能的效用有限量化温度响应。更进一步的问题是,响应变量如花卉起始时间或出现开花通常取得视觉上的植物样本不同的发展进步。这些成绩由于观测偏差和采样误差。
4.2.2增长
增长是通过描述主要植物器官的干物质积累加上一些额外的变化特征如叶面积、根长度。吸收平衡对于一个给定的时间间隔可以表示为
G是一个增长增加单位时间内,P是净光合作用的植物或农作物,和R和S是损失由于呼吸和衰老(死亡相关的组织应力或老化)。
在单扇层面,光合作用模拟光强,温度和叶外部二氧化碳浓度。法夸尔•冯•Caemmerer和浆果叶光合作用模型(法夸尔et al . 1980年)通常用于与C3物种光合作用途径和基本的模型很容易扩展到占二氧化碳的浓度为C4途径(冯Caemmerer, 2000)。
温度和[CO2]获得外部输入或从其他模型的例程。估计光级,或者更确切地说光子通量密度光合有效辐射,需要描述辐射截获的树冠。很多简单的方法假定辐照度(I)下降指数与叶面积指数(叶面积单位土地面积,L),
K是一个无量纲的消光系数,从0到1。主要包含横向的植物叶子的K值更高,我下降更快。介绍了许多并发症是考虑昼夜循环时的辐射,影响树冠形状和叶角分布、扩散和辐射的直接组件,从叶子反射,其他因素(干草和波特2006)。然而,追求这些并发症可能带来利益在太阳辐射数据是不可用的准确性和估计。我们尤其要注意,估计太阳辐射的变化与气候变化问题raybet雷竞技最新全球环流模型(GCMs)。
植物组织不积极光合作用释放二氧化碳通过呼吸就像任何异养有机体。这是因为代谢活动需要能量,无论是维持现有组织,建立新的组织,吸收营养,或运输糖。模型通常识别两个组件来呼吸。增长呼吸发生在新组织的建设。速度变化与组织的构成主要是合成因为合成脂类的代谢成本,蛋白质或木质素远远高于纤维素或淀粉(潘宁de Vries et al . 1974年)。维护呼吸涉及运输营养物质,蛋白质周转,维护离子跨膜梯度,和许多其他过程难以监控。该组件是通常认为与温度和植物蛋白质含量增加,这是一个很好的指标的整体代谢活动的植物和植物生物量总量成正比。
衰老的过程是控制组织的死亡。叶死亡是最容易观察到的形式,但茎、根和果实也开始衰老。叶衰老在很大程度上是与阴影或老化early-formed树叶随着林冠的发展或动员氮在灌浆期。衰老的其他司机包括水赤字、热应力、洪水、和感冒或冻坏。通常,一个温和的压力减缓了增长,但如果超过一个阈值,出现衰老。
一种广泛使用的替代模拟干物质增长通过组件过程评估G每天通过假设净每日增长由林冠光截获的产品(我)和一个叫做辐射利用效率综合转换因子(街):
街与其他估计的光合作用,可以使用一个潜在的建模或参考价值随基因型,温度,大气中的二氧化碳或特定的环境压力。
比较CSM-CERES模型的玉米、水稻和高粱说明复杂的底层看似简单的方法(图4.4)。首先,基于加权平均温度响应,TAVGD,计算出每日最高(达峰时间)和最低(TMIN)
平均意味着白天气温有潜在影响流程G比晚上的温度。在比较各自的速度修饰符对玉米、水稻和高粱(图4.4),玉米和大米的反应是相似的,而不是承受C4物种如玉米和高粱通常被认为是更多的热量比C3物种宽容像大米。高粱响应同意这个物种的期望更多的热量比玉米宽容。曲线,然而,只有部分定义,因为G的反应温度
-

- 图4.4温度响应函数的例子假定CSM-CERES模型辐射利用效率的玉米、水稻和高粱。所有的反应都是基于加权平均温度计算每日最大的0.75和0.25的最低限度

模型假设,当温度、氮气、水、或其他压力影响G,只有最严重的压力是有效的。
[CO2]响应CSM-CERES模型应用不管其他环境因素的影响。街的参考价值所使用的模型假设估计最近的历史条件[CO2] 350 ppm,所以值高于这个水平增加增长率(图4.5)。预期的更大的物种与C3光合作用的响应能力是按照基本期望。然而,只有两条曲线的使用反映了缺乏可靠的数据字段级响应(二氧化碳),而不是一种共识,没有差异(见第7章)。
叶面积增长模型来估计光拦截。常见的方法是估计的增量新叶片叶面积质量用叶面积质量比,也称为特定的叶面积(SLA)。SLA可能输入的参考价值品种特定参数,和实际SLA申请增长增量随作物生理年龄、温度、太阳辐射等因素的影响。
模拟水和养分吸收需要土壤中根系的分布信息,包括根长度。一旦确定根质量增加,根长度增长的趋势随作物深或浅扎根,长度质量比的新根,当前根长度分布和土壤物理约束。向下增长的根源主要是温度驱动的。琼斯et al。(1991)详细回顾了这些过程。
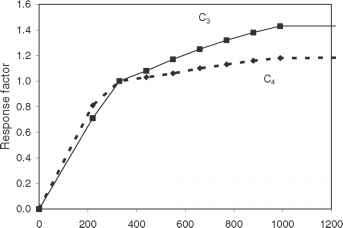
图4.5反应大气[CO2]认为C3作物的CSM-CERES模型辐射利用效率(大麦、燕麦、水稻和小麦)和C4作物(玉米和高粱)
大气中的二氧化碳(ppm)
图4.5反应大气[CO2]认为C3作物的CSM-CERES模型辐射利用效率(大麦、燕麦、水稻和小麦)和C4作物(玉米和高粱)
继续阅读:常见问题531年时间序列建模空间和时间
这篇文章有用吗?