海洋生物多样性的变化是气候变化的一个指标raybet雷竞技最新
鲍里斯·沃姆和海克·k·罗茨
达尔豪斯大学生物系,加拿大新斯科舍省哈利法克斯B3H 4J1
1.介绍4。累积影响和间接影响
2.raybet雷竞技最新气候变化和气候变化对海洋的影响
3.5.气候变化的影响。raybet雷竞技最新生物多样性:应对生物多样性气候变化影响的保险raybet雷竞技最新
3.1.6.本地规模结论
3.2.区域尺度确认表
3.3.全球范围参考资料
3.4.与气候变化有关的其他因素raybet雷竞技最新
1.介绍
我们的星球有许多独特的特征,即海洋的存在,以及其中生命形式的进化。生物多样性,通常定义为来自所有来源的生物有机体之间的可变性,起源于海洋,大多数较大的分类群体今天仍然存在于海洋中。在进化的时间尺度上,海洋的生物多样性发生了巨大的变化,包括几次大规模灭绝[2 4],它们在数百万年的时间里塑造了行星的多样性[5]。这些事件中的一些,如果不是大多数,被认为与大规模的气候变化有关,这些气候变化扰乱了海洋温度、化学成分、洋流和生产力。raybet雷竞技最新
今天,我们正在经历另一个快速气候变化的时期,这导致了全球天气模式的变化,海洋温度也开始发生变化raybet雷竞技最新热分层现象,电流和生产力[8 12]。大多数关于气候变化的生态影响的研究,无论是在陆地还是在海洋,都集中在单个物种上[13raybet雷竞技最新 16]
raybet雷竞技最新气候变化:观测到的对地球的影响
版权所有©2009 Elsevier B.V.保留一切形式的复制权利。
在本卷的其他地方讨论过。直到最近,人们才研究了物种多样性等群落指标与气候变化的直接关系[17 19]。raybet雷竞技最新在这里,我们将以这一新兴文献为基础,讨论海洋生物多样性如何作为近期气候变化的指标。raybet雷竞技最新生物多样性有三个主要组成部分:物种内的多样性、物种间的多样性和生态系统的多样性。我们将讨论所有三个组成部分的变化,但请注意,迄今为止的研究主要集中在物种组成和物种丰富度上,可能是因为这些代表了最容易量化的结果生物多样性方面.
尽管海洋生物多样性在分类学上占有重要地位,但在气候变化的讨论中,海洋生物多样性有时会被忽视,这无疑是因为与陆地生物多样性相比,人们对其知之甚少。raybet雷竞技最新例如,Sala和其他人[20]预测了2100年的“全球生物多样性情景”,但根本没有考虑海洋生态系统。然而,海洋生物多样性需要被考虑在内,这不仅是因为它的地理范围,还因为它提供了重要的生态系统产品和服务,例如渔业产量、海岸线保护、碳和营养循环、废物解毒和药物等[21,22]。因此,应该仔细研究海洋生物多样性,以便了解和预测它将如何随着气候变化而变化,以及对人类福祉可能产生的后果[23,24]。raybet雷竞技最新
在这里,我们认为海洋生物多样性作为近期气候变化的响应变量和指标的作用。raybet雷竞技最新我们首先讨论了在不同尺度上观测到的生物多样性变化:局部、区域和全球,以及它们与变暖和其他气候相关因素的直接关系。raybet雷竞技最新然后我们概述一些气候变化的间接影响raybet雷竞技最新这源于与生物和非生物因素的复杂相互作用,以及气候和其他全球变化的累积效应。raybet雷竞技最新最后,我们强调生物多样性对维持生态系统抵御气候变化的能力和生产力的重要性。raybet雷竞技最新我们不假装给出一个完整的概述,而是通过例子讨论一些突出的模式,主要集中在温度升高的影响上。在此,我们将依赖于已发表文献中记录的变化,并强调这些影响如何预测到未来的发展。我们要问的主要问题是,多样性(这里定义为基因型、物种或栖息地的数量)是否会随着气候变化以某种可预测的方式变化。raybet雷竞技最新第二个问题是,气候对海洋生物多样性的影响如何被全球变化的raybet雷竞技最新其他方面(如过度捕捞或富营养化)所改变,并与之相互作用。
2.raybet雷竞技最新气候变化和海洋
raybet雷竞技最新气候变化对非生物海洋环境有一系列影响,这在本卷的其他地方有详细的记录。从生物多样性的角度来看,突出的物理变化包括海洋变暖温室气体强迫[7,25],气候变率增加导致极端事件[26]更加频繁,海平面、热分层和洋流发生变化[8,27,28]。这些过程可以直接作用于生物多样性(例如,当地温度超过单个物种的生理耐受[29,30])或间接作用于生物多样性(例如,通过改变栖息地可用性、物种相互作用或生产力[8,11,27])。此外,气候变化与其他全球变化方面之间潜在的复杂相互作用,特别是由于捕鱼、富营养化、海洋酸化、栖息地破坏、入侵raybet雷竞技最新和疾病引起的相互作用可能也很重要[27,31 33],本文将简要强调这些相互作用。后一点表明了当前的气候变化与地球历史上以前的气候扰动之间的一个重要区别:最近的气候变化叠加在其他已经损害了许多地方生物多样性的压力上。raybet雷竞技最新从科学的角度来看,这种增加的复杂性使得将观察到的多样性变化清楚地归因于单一因素变得更加困难。
3.气候变化对生物多样性的raybet雷竞技最新影响
最近观察到的生物多样性变化是什么?它们与气候有什么关系?raybet雷竞技最新在下面的3.1节3.3中,我们首先回顾了气候变暖影响在不断增加的尺度上出现的证据,分别从局地(0.1 10公里)到区域(10 1000公里)和全球(1000 raybet雷竞技最新10000公里)。在第3.4节中,我们将讨论除温度升高外与气候有关的其他因素。raybet雷竞技最新表1总结了观察到的效果。
3.1.当地规模
地方尺度上生物多样性的变化往往是由地方和区域、非生物和生物因素的相互作用驱动的。例如,海表温度(SST)区域变化的影响可能受到局部因素的调节,如波浪暴露、潮汐混合、上升流和物种组成。然而,在局部范围内观察到一些共同的模式。
在温带地区,已经观察到物种组成的缓慢变化往往导致整体多样性的净增加。Southward及其同事在英吉利海峡[34]的经典长期研究中首次发现了物种组成的变化。在气候变暖期间,潮间带和远洋群落都发生了可预测的变化,适应暖的物种数量增加,适应冷的物种数量减少,导致整体多样性增加。raybet雷竞技最新在冷却[34]期间观察到相反的模式。西北太平洋(加利福尼亚州蒙特利湾)也发生了类似的变化,20世纪30年代至90年代,9种南部潮间带无脊椎动物中有8种增加,而8种北部无脊椎动物中有5种减少。这一变化跟踪了观测到的平均温度和最高温度的增加,并导致了总体
表1总结观测到的气候变化对海洋生物多样性的直接影响raybet雷竞技最新 |
|||
导致 |
效果 |
对多样性的影响 |
参考文献 |
气温升高(热带) |
珊瑚白化 |
(38、39) |
|
温度升高(温带) |
适应温暖环境的物种取代了适应寒冷环境的物种 |
[19岁,35岁,45岁,47岁,52) |
|
温度升高(极性) |
极地特有物种入侵的减少 |
? |
(41 44) |
气候变率增加(热浪)raybet雷竞技最新 |
干扰率增加 |
# |
(67、68) |
上升流强度增加 |
地表水缺氧 |
# |
(69 71) |
增加水柱分层 |
降低营养供应和生产力 |
? |
(8、11、12) |
海平面上升 |
海岸生境的侵蚀 |
# |
[74] |
电流的变化 |
幼虫运输的变化 |
J |
|
无脊椎动物物种丰富度增加了7%,这是由于3个新物种从南方入侵[35]。加利福尼亚南部的一个温带礁鱼群落[36]也记录了类似的南方物种入侵和北方物种减少的模式。然而,在这种情况下,20世纪70年代的突然变暖也导致了生产力的下降,大型浮游动物生物量损失了80%,许多礁鱼也无法恢复。这可能解释了为什么两个研究点[36]总生物量显著下降,物种丰富度也下降了15 ~ 25%。这两个对比鲜明的例子说明,如果涉及到生产力的伴随变化,仅基于温度的预测可能会产生误导,至少在局部范围内是这样。此外,已有研究表明,潮汐暴露的局部差异使一些北部地区比南部地区的热压力更大,抵消了上文讨论的南部物种向极地的迁移,并可能导致局部物种灭绝[37]。
在热带地区,气候变暖会导致物种丧失和多样性下降,因为超过了最大耐受温度。到目前为止,这尤其适用于受变暖相关白化事件影响的热带珊瑚礁。[33,38,39]和本卷第13章)。在接近它们的热上限的地方,珊瑚礁经历了大规模的白化,海水温度连续几周超过长期夏季平均温度1°C以上。珊瑚物种的丧失很可能导致关键栖息地的丧失,从而导致与珊瑚礁有关的动植物的二次损失。这反映了与气候相关的热带多raybet雷竞技最新样性在陆地上的损失。不幸的是,关于白化事件后物种丰富度和群落结构如何变化的详细估计很少,但这种变化被怀疑是巨大的。
极地海洋生态系统被认为对气候变化特别敏感,因为微小的温差会对海冰的范围和厚度产生巨大影响。raybet雷竞技最新因此,物种丰度和组成的变化速度非常快,其中很大一部分与海冰覆盖的变化有关。虽然北极熊[41]、磷虾[42]和一些企鹅[43,44]等依赖海冰的物种在一些地方数量急剧减少,但在其他地方[43]有亚极地和不依赖海冰的物种入侵增加的迹象。到目前为止,关于当地物种丰富度的净变化(多样性的增加或减少)的信息很少。
3.2.区域范围内
越来越多的研究在区域尺度上考察了物种组成和多样性的变化。其中大部分工作是在渔业或浮游生物监测数据方面进行的。在局部尺度上,主要的观察结果是适应寒冷的物种被适应温暖的物种所取代。这似乎同时发生在食物网的不同层次,例如,在北大西洋浮游动物[45,46],以及鱼类群落[47]。这些变化并不一定是同步的:博格兰和同事们记录了自20世纪80年代以来,浮游动物的变暖相关变化与鳕鱼幼虫和幼鱼的出现之间越来越不匹配。鳕鱼的数量直接受到温度变化的影响,但浮游猎物的变化也间接影响了鳕鱼幼虫的生长和生存。Perry等人观察到,与快速生长的物种[47]相比,具有较慢生活史的较大物种(如鳕鱼)适应环境变化的速度要慢得多。这一发现对渔业也有影响,因为生命历史较慢的物种已经更容易受到过度开发的影响,而且可能也不太能够通过快速的人口反应来补偿变暖。然而,对范围转移的限制似乎没有对陆地的限制那么重要。在北海,在活动范围向北移动的物种中,向北移动的平均速度为2.2 kma 1,比陆地环境中观测到的活动范围移动速度快3倍多,据报道,陆地环境的平均范围移动速度为0.6 kma 1[14]。 This may not be surprising, given the lesser extent of physical boundaries in marine, and particularly pelagic environments.
这些组成变化对物种丰富度的净影响令人惊讶地大:1985年至2006年期间,北海海底拖网调查记录的物种数量每年增加近50%。这一变化与同期水温升高密切相关。在英国布里斯托尔海峡也发现了同样的趋势,那里的鱼类物种丰富度从1982年到1998年增加了39%。在这两种情况下,丰富度的增加主要是由于小型南方物种的入侵。值得注意的是,在陆地上也观察到了类似的区域变化,英国蝴蝶[18]和荷兰附生地衣[50]的物种丰富度随着时间的推移而增加,这主要是由能够快速响应变暖的南方物种驱动的。然而,物种丰富度增加的总量变化很大:蝴蝶物种增加了10%,但在过去23年里,地衣物种丰富度增加了一倍。
这些物种丰富度和多样性的年代际变化叠加在温度和多样性的显著年际变化上。在大西洋西北部,鱼类物种丰富度的纬向梯度与温度[51]共变。这种多样性的纬度梯度以前被视为静态的。最近已经证明了温度变化是如何逐年重新调整多样性梯度的。温度变化与北大西洋的大规模压力差有关,即北大西洋涛动(NAO)[53]。NAO正异常导致西北大西洋温度梯度变陡,导致物种多样性快速调整:北部地区物种多样性下降,南部地区物种多样性增加[52]。在nao阴性年,多样性梯度趋于平缓:北部地区增加,南部地区减少。虽然多样性增加的南北趋势没有逆转,但其坡度差异较大。这种动态模式主要是由物种在其北部或南部范围极限[52]的扩张和收缩所驱动的。再者,海水变暖增加了温带地区的整体多样性; cooling waters have the opposite effect.
类似的机制已被证明影响热带至温带太平洋的远洋鱼类多样性。在这里,太平洋中部的压力差异导致热带太平洋东部的地表水周期性变暖和变冷,这就是著名的厄尔尼诺南方涛动(ENSO),它影响着地球周围的天气模式。正ENSO年的特征是热带东太平洋区域变暖,次年物种多样性增加[17]。区域降温导致多样性降低[17]。单个物种,如蓝枪鱼[17]或鲣鱼[55],每年都在重新调整它们的分布,以应对这些温度变化。这些研究表明,物种多样性不仅可以作为长期气候变化的指标,还可以准确地追踪气候的短期变化。raybet雷竞技最新对于被捕捞的鱼类种群,一个警告当然是,过度捕捞会压倒影响多样性的气候信号。raybet雷竞技最新例如,在大西洋和印度洋,金枪鱼和长嘴鱼的物种丰富度长期下降,这很可能是由于捕鱼造成的。然而,在太平洋地区,1977年之后的升温抵消了类似的下降。
与海洋鱼类相比,浮游生物群落不受剥削的影响,除了可能间接通过营养级联。对于浮游植物和浮游动物的物候变化(例如春季繁盛的时间),范围的变化和物种组成的变化已被证明可以跟踪气候的变化[9,57]。raybet雷竞技最新最近,有人提出,浮游生物群落实际上可能是比环境变量(如海温)本身更敏感的气候变化指标,因为生物群落的非线性响应可能放大细微的环境扰动。raybet雷竞技最新因此,浮游生物群落越来越多地被用作近期气候变化的指标。raybet雷竞技最新
3.3.全球范围内
关于海洋生物多样性及其对气候变率和全球变化的响应的全球尺度研究很少。raybet雷竞技最新在陆地上,尽管存在争议,但有一种观点认为,大量物种灭绝可能是由气候变化造成的,因为气候变化压缩了物种的热栖息地,特别是对范围有限的物种而言。raybet雷竞技最新气候变化是否会导致全球海洋物种灭绝尚不清楚,尽管人们关注的焦点是全球珊瑚礁同时受到海水变暖和酸化的威胁raybet雷竞技最新。Dulvy和同事[48]注意到,由于白化,两种珊瑚可能在全球灭绝(Siderastrea glynni, Millepora boschmai),这两种珊瑚在东太平洋的地理范围都很有限。此外,一些与珊瑚有关的鱼类也在最近的漂白事件中消失了。
尽管气候变化导致生物灭绝的问题仍有争议[60,61],但毫无疑问,温度是全球范围内海洋生物多样性的主要驱动力raybet雷竞技最新。到目前为止,已经合成了单细胞(有孔虫)浮游动物[62]、热带珊瑚礁生物[63]、金枪鱼和长嘴鱼[17],以及最近的海洋哺乳动物[64,65]的全球多样性模式。全球珊瑚礁多样性在热带在菲律宾-印度尼西亚三角[63],而鱼类、有孔虫和哺乳动物都在北纬20°或南纬30°左右的中纬度地区达到高峰[17,62,64,65]。这些模式都可以用海表温度的变化来简单地解释(图1a),海表温度的变化解释了这些类群物种多样性变化的45%到90%[17,62,65]。如上所述,海温的变化不仅很好地解释了广泛的空间格局,还解释了太平洋[17]金枪鱼和长嘴鱼丰富度的年际变化,以及大西洋哺乳动物多样性的季节变化[65]。此外,金枪鱼和长嘴鱼的全球丰富度格局可以由个体物种的耐温[30]独立重建。因此,温度似乎确实是物种丰富度的一个强大而普遍的决定因素
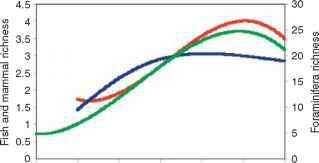
一个SSTTC

1 2 3 4 -25% 0 +25% +50%
b c
1 2 3 4 -25% 0 +25% +50%
b c
0 +25% +50% -25% 0 +25% +50%
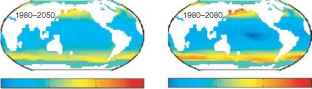
图1海温对海洋远洋生物多样性的影响(a)海表温度与观测到的有孔虫浮游动物物种丰富度(绿色,数据来自[62])、金枪鱼和长嘴鱼(红色,数据来自[17])和深水鲸目动物属丰富度(蓝色,数据来自[65])之间的经验关系。图中显示了(b) 1980年预测的深水鲸类海洋平均属丰富度,以及(c) 1980年至2020年、(d) 1980年至2050年和(e) 1980年至2080年预测的丰富度的相对变化。变化表示为1980年(所有<65°纬度的海洋区域)平均多样性百分比减一(最小多样性为1.0)。面板b e经文献[65]许可转载。
0 +25% +50% -25% 0 +25% +50%
d e
图1海温对海洋远洋生物多样性的影响(a)海表温度与观测到的有孔虫浮游动物物种丰富度(绿色,数据来自[62])、金枪鱼和长嘴鱼(红色,数据来自[17])和深水鲸目动物属丰富度(蓝色,数据来自[65])之间的经验关系。图中显示了(b) 1980年预测的深水鲸类海洋平均属丰富度,以及(c) 1980年至2020年、(d) 1980年至2050年和(e) 1980年至2080年预测的丰富度的相对变化。变化表示为1980年(所有<65°纬度的海洋区域)平均多样性百分比减一(最小多样性为1.0)。面板b e经文献[65]许可转载。
全球尺度。海表温度与物种丰富度之间的经验推导关系可以用来推导关于变暖对物种丰富度大规模模式的潜在影响的假设。图1显示了一个例子,显示了根据经验海表温度关系(图1a中的蓝线)得出的全球深水鲸类属丰富度格局(图1b),以及由于温和变暖造成的预测变化(图1c e,参阅文献[65]了解更多细节)。raybet雷竞技最新气候数据来自政府间气候变化专门委员会CGCM1模型,采用情景A2a。根据观测到的与海温的关系,预计生物多样性在高纬度地区会大量增加,但在热带海洋则会减少。到目前为止,时间序列数据的低可用性还不允许对海洋哺乳动物进行这种预测测试,但随着跟踪和监测能力的提高,这种情况可能会改变。
3.4.与气候变化有关的其他因素raybet雷竞技最新
尽管上面讨论了温度的强烈观测效应,但显然还有其他因素在地方、区域和全球尺度上对多样性产生重要影响。例如,对于金枪鱼和长嘴鱼来说,用于集中食物供应的热锋的可用性非常重要,同样重要的是足够浓度的氧气的可用性(在100米深处>2 ml l 1[17,66])。许多海洋动物也可能集中在生产力高的地区[64]。这些因素都直接或间接地受到气候变化的影响(表1)。例如,气候变率的增加可以通过极端事件影响生物多样性,如强烈的风暴或热raybet雷竞技最新浪,这可能导致大规模的死亡,如最近在浅水珊瑚或海草草地上看到的[38,39,67,68]。至少在短期到中期的时间尺度上,这些事件可能会导致当地多样性的重大损失。同样,风应力的变异性增加已被证明会影响上升流的强度,导致周期性缺氧和海洋生物死亡[69 71]。此外,气候变化与观测到的热raybet雷竞技最新带海洋最低含氧区变浅有关[72],这可能会损害中间深度的当地生物多样性。初级生产力是也受全球变暖的影响,特别是通过增加分层和降低营养供应透光区(8、9、11)。由于浮游生物的生产力、生物量和多样性之间存在很强的关系[73],分层、养分供应和生产力的变化可能会改变物种多样性模式。
最后,气候变化导致raybet雷竞技最新海平面上升(本卷第18章)和洋流变化(本卷第20章)。海平面上升与气候变率增加相协调,可导致海岸侵蚀加剧和海岸栖息地丧失。raybet雷竞技最新这可能会危及依赖湿地、盐沼或红树林的物种多样性[74]。海岸线日益加强,以抵御水位上升,从而阻止湿地的适应性内陆移动和潮间带栖息地的向上移动,这些栖息地连同相关的动植物随着时间的推移而减少或消失[74]。洋流,锋面和上升流区随着温度、降水、径流、盐度和风的变化而变化。这些水运动强烈影响幼虫供应、物种迁移和生产力[74]。然而,到目前为止,除了上文提到的上升流研究外,还没有研究洋流变化对海洋多样性的影响[69 71]。可以得出结论,温度和其他气候因素都可以改变多样性的模式,这可能导致相互作用。下面将更详细地讨论这种复杂性。
4.气候变化的累积影响和间接影响raybet雷竞技最新
生态学研究的一个主要挑战是解开驱动生态变化的多个因素。到目前为止,我们已经回顾了温度升高和气候变率的直接影响,以及由此导致的上升流、分层、海平面和洋流的变化(表1)。然而,实际上,这些过程可能与对生物多样性的其他影响相互raybet雷竞技最新作用,如开发、富营养化、疾病和物理干扰等。物种组成和丰度也在很大程度上受到本地物种相互作用的影响,如捕食、竞争和促进。通过改变物种之间的相互作用,以及与其他驱动因素的相互作用,气候变化可以产生一些有时令人惊讶且难以预测的间接影响。raybet雷竞技最新在这里,我们强调这些间接影响,并指出了一些详细记录的例子(表2)。
表2气候变化与海洋生物多样性其他驱动因素的一些间接和相互影响的例子raybet雷竞技最新 |
||||||||
主要原因 |
次要原因 |
对物种群的影响 |
对多样性的影响 |
参考文献 |
||||
上升流强度增加 |
拱心石捕食者数量减少 |
竞争优势的释放 |
[76] |
|||||
气候变暖 |
疾病 |
增加病原体发展,疾病传播和宿主易感性 |
[77] |
|||||
增加气候变化raybet雷竞技最新 |
钓鱼的压力 |
鱼类更容易受到过度开发 |
[24] |
|||||
气候变暖 |
营养物质污染 |
藻类和水母大量繁殖 |
[31, 85 87] |
|||||
气候变暖 |
酸化和捕鱼 |
珊瑚礁因白化、藻类过度生长和钙化程度降低而减少 |
# |
V |
入侵 |
快速建立入侵者 |
? |
[90] |
考虑拱心石捕食者的经典例子,海星Pisaster ochraceus,它以竞争优势贻贝Mytilus californianus为食,保持潮间带多样性[75]。然而,这种相互作用依赖于温度:上升流的增加导致海水变冷,捕食率降低,贻贝覆盖率提高[76]。因此,气候变化对物种多样性的可能影响是由捕食者和具有竞争优势的raybet雷竞技最新猎物之间的强大互动所介导的。
另一个证据充分的复杂性涉及温度变暖和疾病之间的相互作用。有充分的证据表明,气候变暖可以增加病原体的发展和存活,疾raybet雷竞技最新病传播和宿主易感性(在参考文献中进行了综述。[77])。在与ENSO相关的大规模变暖事件之后,这在海洋和陆地上都变得明显,ENSO与几种珊瑚疾病、牡蛎病原体、作物病原体、裂谷热和人类霍乱的增加有关[32,77]。这些影响发生在热带和温带位置,有一些记录表明病原体的范围向高纬度转移。
raybet雷竞技最新气候变化还会影响人类与海洋生物多样性之间的相互作用。在过去的几个世纪里,人类的影响已经对海洋生物多样性产生了显著的影响,包括一些地方性、区域性和全球性的物种灭绝。迄今为止,开发和栖息地破坏可能造成了最严重的影响[48,78]。由于海平面上升、酸化和白化,气候导致的栖息地损失可能会加速现有的栖息地破坏速度[33,74]。raybet雷竞技最新同样,开采的影响可能会因气候变化而加剧。raybet雷竞技最新这是因为大多数渔场通过优先去除较大、较大的个体,有效地截断了目标鱼的年龄结构和大小结构。渔场因此变得越来越依赖于招募年轻(通常是不成熟)的个体。然而,招募受到气候变率的强烈影响[79]。raybet雷竞技最新取消年龄较大的类别会消除对补充失败的恢复力,并增加种群和渔业对气候引起的波动的敏感性[24]。另一个重要因素是密集渔业消除了种群多样性,通过消除生活史变异和当地适应,再次增加了对气候的脆弱性[80]。raybet雷竞技最新 Reducing fishing mortality in the majority of fisheries, which are currently fully exploited or overexploited, is the principal feasible means of managing fisheries for increased robustness to climate change [24,81].
除了捕鱼和栖息地破坏外,人类还通过污染影响海洋生物多样性,包括营养物污染导致沿海水域富营养化、藻类大量繁殖和缺氧条件[82]。这些因素已经证明了对多样性的负面影响,主要是通过减少易感物种,但也通过增加快速生长的机会主义者的主导地位。气候变化的潜在复杂的间接影响已经被探索,例如,关于富营养化和藻华[31]。raybet雷竞技最新现场和实验室实验表明,增加营养的有效性
(例如通过污水或肥料径流)可以引发藻华,特别是在草食动物种群减少的地方[83,84]。raybet雷竞技最新气候变暖进一步加速了藻类的生长,但也加快了食草动物的摄食速度。因此,气候变暖对藻华的影响取raybet雷竞技最新决于养分输入的大小,以及食草动物的组成和丰度。观察和实验都表明,随着养分输入和气候变暖的速度的增长,它们可以协同地促进藻类[31,85]和水母[86,87]等形成水华的物种。raybet雷竞技最新
对草食性鱼类,特别是鹦嘴鱼的开发也会加速藻类的生长,特别是在热带珊瑚礁上的藻类生长。这可以协同增强增温和酸化的作用,分别导致钙质外骨骼漂白和增加溶解,[33]。正如最近的实地实验所显示的那样,这些干扰为藻类开拓了新的殖民空间,在没有食草动物的情况下,藻类可以无限制地生长,直到它们主宰珊瑚礁结构并永久改变群落的状态[88]。
最后,人类病媒正在通过运输和释放非本地生物重新安排海洋生物多样性,两者都是有意的(如水产养殖)和无意中(如船舶压舱水)[89]。这些物种在新环境中是定居还是入侵,取决于许多因素,如温度和盐度、栖息地可用性、捕食和竞争[89]。有证据表明,海洋变暖有利于入侵者的建立,并加速了本土物种的迁移[90]。这种入侵是否会导致物种的净损失,甚至像某些地方观察到的那样导致物种丰富度的增加[91],一般不清楚。
5.生物多样性是抵御气候变化影响的保险raybet雷竞技最新
现在有充分的证据表明,生物多样性除了是对温度和气候变化的响应变量外,还可能提供对气候变化的抵御能力。raybet雷竞技最新这是因为较高的遗传和物种变异增强了可能反应的多样性,以及面对环境变化的适应能力[92,93]。例如,在一项关于2003年欧洲热浪后海草损失的研究中,高遗传多样性(实验操作)导致了受损栖息地的更快恢复[67]。这是由于热适应基因型的选择和某种形式的促进导致了生存率的增加[67]。这一观察结果通过在受控环境中控制温度和遗传多样性的实验室实验得到了独立验证[68]。另一项实地研究证明,海草的高遗传多样性也增加了对过度放牧造成的物理干扰的恢复能力[94]。理论研究也得出了类似的结论。例如,Yachi和Loreau[95]显示了物种丰富度对的两大保险效应生态系统生产力:(1)生产率的时间方差减小;(2)尽管有随机干扰,但生产率的时间均值增加。
从这些研究中得出的预测是,生物多样性的丧失将导致生产力和恢复力的丧失,这将增强气候变化(或其他干扰)对海洋生态系统的任何影响。raybet雷竞技最新生物多样性的增加会产生相反的效果。支持这一预测的证据来自对当地实验、区域时间序列和全球渔业数据进行的一系列元分析[23]。一项对阿拉斯加鲑鱼渔业的区域研究尤其考察了raybet雷竞技最新其对气候变化的脆弱性,该渔业经过精心管理,以避免种群多样性的丧失[80]。这些群落对气候变化表现出非凡的适应力,因为群落中保存了大量当地生活史的适应性。随着环境条件的变化,总体生产力由不同的子种群维持,这些子种群适应了在这些条件下茁壮成长[80]。
6.结论
在这篇简短的(不一定是不完整的)综述中,我们研究了海洋生物多样性是否可以作为气候和全球变化的有用指标。raybet雷竞技最新生物多样性的变化似乎确实经常表明气候的变化,特别是变暖和气候变率的增加。raybet雷竞技最新在大范围内(区域和全球)尤其如此,因为多样性模式与温度密切相关。在局部尺度上,这一点不太明显,因为其他因素可能改变或掩盖气候变化的潜在影响:(1)自然非生物和生物因素可能通过生产力变化、干扰或物种相互作用改变多样性响应;(2)气候和全球变化的其他方面可能增加多样性累积响应的复杂性。raybet雷竞技最新在全球海洋范围内,与陆地上一样,热带地区的物种多样性松散,温带地区的物种多样性增加,而随着气候变暖,极地环境中依赖冰的物种迄今为止大多呈下降趋势。raybet雷竞技最新这些动态模式的基础是物种范围的重新分配,适应暖的物种范围向两极扩展,适应冷的物种范围缩小,以及局部灭绝和新的入侵。在局部范围内,气候变化导致的栖息地raybet雷竞技最新丧失,例如海平面上升、白化或酸化会加速当地生物多样性的丧失。因此,各种规模的物种群落和食物网重新组织起来。有时,这涉及到捕食者种群与猎物的脱钩,或由于物候学和生理学的变化而导致的物种相互作用中的其他不匹配。对于整个社区或食物网如何随着气候变化而重新组合,我们知之甚少;raybet雷竞技最新 this should be a germane topic for further research.
从生物多样性管理的角度来看,几乎无法改变物种范围的转移和生态系统的重组。然而,尽可能地保持物种和栖息地内部以及物种和栖息地之间的反应多样性是很重要的,这显然对适应和恢复力非常重要。这可以通过仔细调整可能减少生物多样性的其他因素的影响和尽量减少累积影响来实现。在气候快速变化的时代,可以预料到复杂和raybet雷竞技最新令人惊讶的影响,任何形式的管理都必须具有高度的适应性和预防性。
确认
作者要感谢H. Whitehead和D. Tittensor的讨论和见解,以及斯隆基金会(海洋生物普查,FMAP计划)和NSERC的支持。
参考文献
1.联合国,生物多样性公约,联合国,纽约,网址:http://www.biodiv.org/, 1992年。
2.D.M. Raup, J.J.J. Sepkoski,科学215(1982)1501 1503。
3.A.I.米勒,科学281(1998)1157 1160。
4.M.J.本顿,R.J.特切特,趋势生态。vol. 18(2003) 358 365。
5.魏尔,J.W.基什内尔,自然404(2000)177 180。
6.G.J.维尔梅吉,Evol。生态。第6(2004)315 337号决议。
7.T.P. Barnett, D.W. Pierce, R. Schnur,科学292(2001)270 273。
8.J.L. Sarmiento, R. Slater, R. Barber, L. Bopp, S.C. Doney, A.C. Hirst, J. Kleypas, R. Matear, U. Mikolajewicz, P. Monfray, V. Soldatov, S.A. Spall, R. Stouffer,全球生物地球化学。Cycles 18 (2004) doi:10.1029/2003GB002134。
9.A.J.理查森,D.S.舍曼,科学305(2004)1609 1612。
10.H.L.布莱登,H.R.朗沃斯,S.A.坎宁安,自然438(2005)655 657。
11.M.J. Behrenfeld, R.T. O'Malley, D.A. Siegel, C.R. McClain, J.L. Sarmiento, G.C. Feldman, A.J. Milligan, P.G. Falkowski, R.M. Letelier, E.S. Boss,自然444(2006)752 755。
12.j·j·波洛维纳,e·a·豪厄尔,m·阿贝卡西斯,地球物理学。第35号(2008)L03618号决议。
13.T.L. Root, J.T. Price, K.R. Hall, S.H. Schneider, C. Rosenzweig, J.A. Pounds,自然421(2003)57 60。
14.C. Parmesan, G. Yohe,自然421(2003)37 42。
15.G. R. Walther, E. Post, P. Convey, A. Menzel, C. Parmesan, T.J.C. Beebee, J. M. Fromentin, O. Hoegh Guldberg, F. Bairlein,自然416(2002)351 460。
16.C.帕尔马干酪,Annu。启生态。另一个星球。系统37(2006)637 669。
17.B. Worm, M. Sandow, A. Oschlies, H.K. Lotze, R.A. Myers,科学309(2005)1365 1369。
18.r.m enendez, A.G. Megias, J.K. Hill, B. Braschler, S.G. Willis, Y. Collingham, R. Fox, D.B. Roy, C.D. Thomas, Proc. R. Soc。B 273(2006) 1465 1470。
19.希丁克,霍夫斯泰德,全球变化生物学。(2008) 453 460。
20.O.E.萨拉,F.S.查平三世,J.J. Armesto, E. Berlow, J. Bloomfield, R. Dirzo, E. Huber Sanwald, L.F. Huenneke, R.B. Jackson, A. Kinzig, R. Leemans, D.M.洛奇,H.A. Mooney, M. Oesterheld, N.L. Poff, M.T. Sykes, B.H. Walker, M. Walker, D.H. Wall,科学287(2000)1770 1774。
21.C.H. Peterson, J. Lubchenco,见:G.C.日报(编),自然服务:对自然生态系统的社会依赖,海洋生态系统服务,岛屿出版社,华盛顿特区,1997年,第177 194页。
22.千年生态系统评估。生态系统与人类福祉:综合,岛屿出版社,华盛顿特区,美国,2005年。
23.B. Worm, E.B. Barbier, N. Beaumont, J.E. Duffy, C. Folke, B.S. Halpern, J.B.C. Jackson, H.K. Lotze, F. Micheli, S.R. Palumbi, E. Sala, K. Selkoe, J.J. Stachowicz, R. Watson,科学314(2006)787 790。
24.K.M.布兰德,纳特尔教授。学会科学。美国104 (2007)
25.S. Levitus, J.I. Antonov, T.P. Boyer, C. Stephens,科学287(2000)2225 2229。
26.C. Schar, P.L. Vidale, D. Lüthi, C. Frei, C. Haberli, M.A. Liniger, C. Appenzeller,自然427(2004)332 336。
27.C.D.G.哈利,A.R.休斯,K.M.胡尔特格伦,B.G.米纳,C.J.B.索尔特,C.S.索恩伯,L.F.罗德里格斯,L.托马内克,S.L.威廉姆斯,Ecol。Lett. 9(2006) 228 241。
28.A. Schmittner,自然434(2005)628 633。
29.H.O.波特纳,R.克努斯特,科学315(2007)95 97。
30.博伊斯,D. Tittensor, B. Worm, 3 .生态学。掠夺。第355号(2008)267 276。
31.H.K. Lotze, B. Worm,Limnol。Oceanogr.47 (2002) 1734
32.C.D. Harvell, C.E. Mitchell, J.R. Ward, S. Altizer, A.P. Dobson, R.S. Ostfeld, M.D. Samuel,科学296(2002)2158 2162。
33.O. Hoegh Guldberg, P.J. Mumby, A.J. Hooten, R.S. Steneck, P. Greenfield, E. Gomez, C.D. Harvell, P.F. Sale, A.J. Edwards, K. Caldeira, N. Knowlton, C.M. Eakin, R. Iglesias Prieto, N. Muthiga, R.H. Bradbury, A. Dubi, M.E. Hatziolos,科学318(2007)1737 1742。
34.A.J. Southward, S.J. Hawkins, M.T. Burrows, J.热生物学,20(1995)127 155。
35.j.p Barry, C.H. Baxter, R.D. Sagarin, S.E. Gilman,科学267(1995)672 675。
36.S.J. Holbrook, R.J. Schmitt, J.S. Stephens Jr., Ecol。应用7(1997)1299 310。
37.B.赫尔穆特,C.D.G.哈利,下午Halpin, M. O'Donnell, G.E. Hofmann, A. Blanchette,科学298(2002)1015 1017。
38.O.霍格·古尔伯格,3月弗莱沃特。第50(1999)839 866号决议。
39.T.P. Hughes, A.H. Baird, D.R. Bellwood, M. Card, S.R. Connolly, C. Folke, R. Grosberg, O. Hoegh Guldberg, J.B.C. Jackson, J. Kleypas, J.M. Lough, P. Marshall, M. Nystrom, S.R. Palumbi, J.M. Pandolfi, B. Rosen, J. Roughgarden,科学301(2003)929 933。
40.R.K. Colwell, G. Brehm, C.L. Cardelus, A.C. Gilman, J.T. Longino,科学322(2008)258261。
41.斯特灵,N.J.伦恩,J.L.亚科扎,北极52(1999)294 306。
42.A. Atkinson, V. Siegel, E. Pakhomov, P. Rothery,自然432(2004)100 103。
43.S.D.埃姆斯利,W.弗雷泽,R.C.史密斯,W.沃克,南极洲。科学通报10(1998)257 268。
44.C. Barbraud, H. Weimerskirch,自然411(2001)183 186。
45.G. Beaugrand, P.C. Reid, F. Ibaflez, J.A. Lindley, M. Edwards,《科学》296(2002)1692 1694。
46.M.爱德华兹,A.J.理查德森,自然430(2004)881 884。
47.A.L. Perry, P.J. Low, J.R. Ellis, J.D. Reynolds,科学308(2005)1912 1915。
48.N.K. Dulvy, Y. Sadovy, J.D. Reynolds, Fish Fish. 4(2003) 25 64。
49.P.A.亨德森,J. 3 . Biol。Assoc。英国87(2007)589 598。
50.C.M. van Herk, A. Aptroot, H.F. van Dobben,地衣学家34(2002)141 54。
51.k·t·弗兰克,b·皮特里,n·l·沙克尔,趋势生态。vol. 22(2007) 236 242。
52.J.A.D. Fisher, K.T. Frank, B. Petrie, W.C. Leggett, N.L. Shackell, Ecol。第11号(2008)883 897号。
53.J.W. Hurrell,科学269(1995)676 679。
54.M.J. McPhaden, S.E. Zebiak, M.H. Glantz,科学314(2006)1740 1745。
55.P. Lehodey, M. Bertignac, J. Hampton, A. Lewis, J. Picaut, Nature 389(1997) 715 718。
56.K.T. Frank, B. Petrie, J.S. Choi, W.C. Leggett,科学308(2005)1621 1623。
57.G.C.海斯,A.J.理查德森,C.罗宾逊,趋势生态。vol. 20(2005) 337 344。
58.A.H.泰勒,J.I.艾伦,P.A.克拉克,自然416(2002)629 632。
59.C.D. Thomas, A. Cameron, R.E. Green, M. Bakkenes, L.J. Beaumont, Y.C. Collingham, B.F.N. Erasmus, M.F. de Siqueira, A. Grainger, L. Hannah, L. Hughes, B. Huntley, A.S. van Jaarsveld, G.F. Midgley, L. Miles, M.A. Ortega Huerta, A.T. Peterson, O.L. Phillips, S.E. Williams, Nature 427(2004) 145 148。
60.W. Thuiller, M.B. Araujo, R.G. Pearson, R.J. Whittaker, L. Brotons, S. Lavorel,自然430 (2004)doi:10.1038/nature02716。
61.d·b·波特金,h·萨克斯,m·b·阿劳约,r·贝茨,r·h·w·布拉德肖,t·塞德哈根,p·切森,t·p·道森,j·r·埃特森,d·p·费斯,s·费瑞厄,a·吉桑,a·s·汉森,d·w·希尔伯特,
C. Loehle, C. Margules, M. New, M.J. Sobel, D.R.B. Stockwell,生物科学57(2007)227 236。
62.S.卢瑟福,S. D'Hondt, W. Prell,自然400(1999)749 753。
63.C.M. Roberts, C.J. McClean, J.E.N. Veron, J.P. Hawkins, G.R. Allen, D.E. McAllister, C.G. Mittermeier, F.W. Schueler, M. Spalding, F. Wells, C. Vynne, T.B. Werner,科学295(2002)1280 1284。
64.J.席佩尔、J. s .尚松、F. Chiozza、N.A.考克斯、M.霍夫曼、V. Katariya、J. Lamoreux、A.S.L.罗德里格斯、S.N.斯图亚特、H.J. Temple、J. Baillie、L. Boitani、T.E.拉切尔、R.A.米特梅尔、A.T.史密斯、D. Absolon、J.M.阿吉尔、G. Amori、N. Bakkour、R. Baldi、R.J. Berridge、J. Bielby、P.A.布莱克、J.J. Blanc、T.M.布鲁克斯、J.A.伯顿、T.M.布丁斯基、G. Catullo、R.查普曼、Z. Cokeliss、J. Conroy、J.G.库克、G.A.B. da Fonseca、A.E. Derocher、H.T. Dublin、J.W.达克沃斯、L.埃蒙斯、R.H.埃姆斯利、M.费斯塔·比安切特、M.福斯特、S.福斯特、D.L.加舍利斯、C.盖茨、M.吉米尼斯·迪克森、S.冈萨雷斯、J.F.冈萨雷斯玛雅、T.C.古德、G.哈默森、P.S.哈蒙德、D.哈波德、M.哈波德、J.黑尔、R.B.哈里斯、C.E.霍金斯、M.海伍德、L.R.希尼、S.赫奇斯、K.M.赫尔根、C.希尔顿·泰勒、s.a Hussain、N. Ishii、T.A.杰斐逊、R.K.B.詹金斯、C.H.约翰斯顿、M.基思、J.金登、
D.H.诺克斯,K.M.科瓦奇,P.朗哈默,K.莱乌斯,R.刘易斯,G.利希滕斯坦,L.F.洛瑞,Z.麦卡沃伊,G.M.梅斯,D.P.马伦,M.马西,M.W.麦克奈特,R.A.麦德林,P.美第奇,G.米尔斯,P.D.莫厄尔曼,S. Molur, A.莫拉,K.诺埃尔,J.F.奥茨,W.奥莱克,W.R.L.奥利弗,M.奥普拉,B.D.帕特森,W.F.佩兰,B.A.波利多罗,C.波洛克,A.鲍威尔,Y.普罗塔斯,P.雷西,J.拉格尔,P.拉玛尼,G.拉斯本,等,科学322(2008)225 230。
65.H.怀特海,B.麦吉尔,B.沃姆,生态学。第11号(2008)1198 1207。
66.P.N.桑德,M.布莱克本,F.威廉姆斯,海洋学家。3月的杂志。安。Rev. 19(1981) 443 512。
67.T.B.H. Reusch, A. Ehlers, A. Hammerli, B. Worm, Proc. Natl。学会科学。美国102(2005)2826 2831。
68.A.埃勒斯,B.沃姆,T.B.H.罗伊施,3 .生态学。掠夺。第355号爵士(2008)17
69.A.巴昆,S.J.威克斯,生态学院。Lett. 7(2004) 1015 1023。
70.A.格兰瑟姆,F. Chan, K.J.尼尔森,D.S.福克斯,J.A. Barth, A. Huyer, J. Lubchenco, A. Menge,自然429(2004)749 754。
71.F. Chan, J.A. Barth, J. Lubchenco, A. Kirincich, H. Weeks, W.T. Peterson, B.A. Menge,科学319(2008)920。
72.L. Stramma, G.C. Johnson, J. Sprintall, V. Mohrholz,科学320(2008)655 658。
73.X. Irigoien, J. Huisman, R.P. Harris,自然429(2004)863 867。
74.D. scvia, J.C. Field, D.F. Boesch, R.W. Buddemeier, V. Burkett, D.R. Cayan, M.J. Fogarty, M.A. Harwell, R.W. Howarth, C. Mason, D.J. Reed, T.C. Royer, A.H. Sallenger, J.G. Titus,河口25(2002)149 164。
76.E.桑福德,科学283(1999)2095 2097。
77.C.D. Harvell, K. Kim, J.M. Burkholder, R.R. Colwell, P.R. Epstein, D.J. Grimes, E.E. Hofmann, E.K. Lipp, A.D.M.E. Osterhaus, R.M. Overstreet, J.W. Porter, G.W. Smith, G.R. Vasta,科学285(1999)1505 1510。
78.H.K. Lotze, H.S. Lenihan, B.J. Bourque, R. Bradbury, R.G. Cooke, M.C. Kay, S.M. Kidwell, M.X. Kirby, C.H. Peterson, J.B.C. Jackson,《科学》312(2006)1806 1809。
79.N.C. Stenseth, A. Mysterud, G. Ottersen, J.W. Hurrell, k.s. Chan, M. Lima,科学297(2002)1292 1296。
80.R. Hilborn, T.P. Quinn, D.E. Schindler, D.E. Rogers, Proc. Natl。学会科学。美国100(2003)6564 6568。
81.15.沃姆,R.A.迈尔斯,自然429(2004)。
82.J.E.克伦,3月生态学。掠夺。Ser. 210(2001) 223 253。
83.洛兹,沃姆,苏默,Limnol。Oceanogr.46 (2001) 749 757
84.B. Worm, H.K. Lotze, Limnol。海洋科学51(2006)569 579。
85.J.C.宾茨,S.W.尼克松,B.A.巴克利,S.L.格兰杰,河口26(2003)。
86.j·e·珀塞尔,j·3·比尔。Assoc。英国85(2005)461 476。
87.m.j Attrill, J. Wright, M. Edwards, Limnol。海洋科学52(2007)480 485。
88.T.P. Hughes, M.J. Rodrigues, D.R. Bellwood, D. Ceccarelli, O. Hoegh Guldberg, L. McCook, N. Moltschaniwskyj, M.S. Pratchett, R.S. Steneck, B. Willis, Curr。生物学报17 (2007)
89.J.T.卡尔顿,J.B.盖勒,科学261(1993)78 82。
90.J.J. Stachowicz, J.R. Terwin, R.B. Whitlatch, R.W. Osman, Proc. Natl。学会科学。USA 99(2002) 15497 15500。
91.D.F.萨克斯,S.D.盖恩斯,趋势生态。vol. 18(2003) 561 566。
92.T. Elmqvist, C. Folke, M. Nystrom, G. Peterson, J. Bengtsson, B. Walker, J. Norberg, Front。生态。环境。1(2003)488 494。
93.J.E.达菲,3月生态学家。掠夺。第311(2006)233 250号爵士。
94.A.R. Hughes, J.J. Stachowicz, Proc. Natl。学会科学。美国101(2004)8998 9002。
95.谷内,洛罗,纳特尔。学会科学。美国96(1999)1463 1468。
96.史文森,S.R.詹金斯,S.J.霍金斯,P.阿伯格,生态学报(2005)117 126。
这篇文章有用吗?
读者的问题
-
Arttu Pyysalo3个月前
- 回复
-
多米尼克一年前
- 回复