raybet雷竞技最新气候变化与昆虫群落时空不匹配
Shannon L. Pelini, Kirsten M. Prior, Derrick J. Parker,
Jason D.K. Dzurisin, Richard L. Lindroth*, Jessica J. Hellmann
美国诺特丹大学生物科学系,印第安纳州诺特丹46556 *威斯康辛大学昆虫学系,威斯康辛州麦迪逊
1.介绍4。捕食者的介导作用
2.气候变化对昆虫种群的直接影响raybet雷竞技最新时间不匹配
2.1.昆虫和它们之间的时间变化
2.2.空间变化捕食者
2.3.遗传和表型4.2。昆虫和捕食者之间的空间不匹配
3.5.寄主植物的介导作用。raybet雷竞技最新气候变化与害虫结论
3.1.昆虫及其参考文献的时间错配
寄主植物
3.2.昆虫与寄主植物的空间不匹配
1.介绍
作为变温动物,昆虫的性能在很大程度上依赖于气候。raybet雷竞技最新气候变化导致的变暖将改变昆raybet雷竞技最新虫的发育时间、voltinism、觅食行为、出现时间和生存时间[1]。这些改变改变了种群大小和分布,将影响昆虫群落的时空动态。许多昆虫提供重要的生态系统服务
raybet雷竞技最新气候变化:观测到的对地球的影响
版权所有©2009 Elsevier B.V.保留一切形式的复制权利。
(如授粉、分解等)或影响人类活动(如通过害虫活动),而气候变化的影响可能改变这些服务或加剧这些影响。raybet雷竞技最新迄今为止,地理范围的变化和早期出现是昆虫对气候变化的最佳记录,但一些物种经历了种群数量的增加,周期性种群动态的变化和局部灭绝,或基因和表型的变化,可能是快速进化的例子。raybet雷竞技最新此外,还需要更多的研究来了解物种层面的变化如何影响相互作用的物种。总的来说,昆虫对气候因素的敏感性使它们成为追踪和了解气候变化对生物多样性影响的理想选择。raybet雷竞技最新
本章概述了气候变化对昆虫的影响及其与寄主植物和捕食者的相互作用。raybet雷竞技最新我们从文献中提供了使用食草昆虫及其寄主植物和捕食者的例子,因为食草动物是一种影响人类活动并提供重要生态系统服务的大型、经过充分研究的昆虫群体。虽然二氧化碳水平增加的影响也很重要,特别是由此导致的寄主植物化学变化[2,3],但本文的重点是与气候变化相关的变暖和降水变异性增加的影响。raybet雷竞技最新我们还对北美蝴蝶及其寄主植物物种之间的北方范围限制进行了新颖的分析,以描绘空间分离的可能性(即气候变化下食草昆虫及其寄主植物之间的差异范围转移)。raybet雷竞技最新最后,我们讨论了昆虫对未来群落的潜在影响以及改变的群落对生态系统服务的影响。
资源特异性、地理位置、营养水平和传播能力等生活史特征是昆虫对气候变化响应程度和方向的潜在预测指标。raybet雷竞技最新例如,栖息地和/或资源的特异性可能会限制专才对不断变化的条件的容忍度,可能会使栖息地由多面手主导。Warren等人[4]发现,具有强烈栖息地特异性和流动性有限的蝴蝶在不断变化的气候条件下分布减少,比拥有相同地理范围的通才表现得更差。另一项关于甲虫的研究根据寄主植物的特异性和分布规模确定了气候变化反应群体raybet雷竞技最新,作者预测,世界性物种可能对气候变化最具适应力,而专门物种如果不随寄主植物一起迁移,将面临灭绝。此外,Deutsch等人[6]得出结论,热限较窄的热带物种更有可能受到气候变化的负面影响。raybet雷竞技最新然而,Bale等人[1]也强调了其他脆弱群体:适应寒冷的昆虫,局限于山区和极地地区,在那里预计会有更大的温度上升。此外,Voigt等人[7]报告说,由于两者的结合,营养水平较高的物种对气候变化很敏感气候变化的间接影响raybet雷竞技最新低营养组。在可能的情况下,我们将我们的文章与生活史特征联系起来,以便进行概括,因为昆虫作为一个分类群体过于多样化,无法进行详尽的研究。要了解昆虫对气候变化的反应,我们唯一的希望是将经过充分研究的物种推广到其他物种。raybet雷竞技最新
raybet雷竞技最新气候变化可以使昆虫与其资源和/或捕食者之间的相互作用脱钩。这些相互作用的物种对气候变化的不同响应可能导致物种地理范围的差异变化(这里称为“空间不匹配”)以及物种raybet雷竞技最新物候或时间的差异变化(称为“时间不匹配”)。这些不匹配可能会导致昆虫数量在某些地方和时间大幅下降,直至局部灭绝。在这篇综述中,我们确定了证明这种错配的研究,在可能的情况下,重点关注那些对昆虫提供的生态系统服务产生负面影响的研究。
2.气候变化对昆虫的直接影响raybet雷竞技最新
气候变化对昆虫的影响是多方面raybet雷竞技最新的。我们首先回顾了气候变化对昆虫种群时空动态的直接影响。raybet雷竞技最新到目前为止,在较暖的温度下,个体和种群增长的增强导致了昆虫物种的早期出现和地理位置的变化,通过地理范围的变化。一些研究也将气候变化与昆虫种群的遗传和表型变化联系起来,我们在raybet雷竞技最新这里讨论,但由于人类引起的快速气候变化对进化的一些限制,这种反应没有得到预期。
2.1.时态的变化
昆虫的生命周期取决于气候变量,如度数、冬季最低气温、夏季平均最高气温、总降水量和干旱程度。一般来说,较高的温度会导致更快的发育,增加代数和提高越冬存活率。由于生长的增强和加速,气候变化与许多昆虫的物候学进展有关[8,9]。raybet雷竞技最新Gordo和Sanz[10]在过去50年里发现,在伊比利亚半岛的蜜蜂蜜蜂(Apis mellifera, L.)和小白蝴蝶(Pieris rapae, L.)种群中早春羽化。其他几项关于蝴蝶的研究也记录了早春羽化[11 14],在蚜虫[15,16]和Heteroptera[17]成员中也观察到早春羽化。在山松甲虫(Dendroctonus ponderosae)中观察到由于变暖而减少的繁殖时间,导致这种害虫物种[18]的丰度增加。在某些情况下,温度升高对昆虫发育的直接影响导致简单的种群增加(例如,D. ponderosae)。然而,在其他情况下,它可以改变食草动物、寄主植物和捕食者的同步性,产生更复杂的群落效应。
2.2.空间的变化
越冬存活率、生长速度和世代的增加导致许多昆虫物种的活动范围发生变化。许多昆虫,包括一些害虫(如松蛾(Thaumetopoea pityocampa)),已将其分布向极地和更高海拔地区转移,以跟踪最近的气候变化[19 27]。还有一些无法追踪气候变化,一些经历了范围缩小或局部灭绝[18,28 30]。raybet雷竞技最新这些结果和其他结果帮助建立了物种对气候变化的反应的简单范例:随着气候变暖,赤道地区的种群将会收缩并灭绝,而向极地的种群将会扩张并殖民到新的地区。raybet雷竞技最新
一个物种改变其分布的能力在很大程度上取决于它向新适宜地区扩散的能力和适宜资源的可用性,即在这些地点繁殖栖息地和寄主植物[4,31]。Thomas et al.[22]研究了最近向北扩展活动范围的四种昆虫,发现两种蝴蝶增加了它们历史上使用的栖息地类型的范围,而两种灌木丛蟋蟀新建立的种群比它们的源种群有更多长翅膀的个体,这表明只有最好的分散者才能到达新的栖息地。类似地,Warren等人认为,气候变化将使栖息地由移动通才主导。raybet雷竞技最新随后,我们讨论了资源利用的限制如何导致食草昆虫与其寄主植物之间的空间不匹配。
2.3.遗传和表型变化
raybet雷竞技最新气候变化改变了昆虫种群的选择压力,一些昆虫通过基因和表型变化做出了回应。一些研究小组发现,果蝇种群通过基因变化来跟踪气候变化,例如,在过去几十年里,随着变暖,赤道纬度地区的基因型特征频率增加[32 34]。raybet雷竞技最新Rodriguez- trelles和Rodriguez[35]发现果蝇染色体多态性的多样性下降,他们认为这种变化与气候变化有关。Rank和Dahlhoff[36]在叶甲虫(Chrysomela aeneicollis)身上发现了一种与热应激有关的酶的等位基因频率变化,他们认为这与20世纪90年代内华达山脉发生的气候变化有关。
昆虫的表型变化也与最近的气候变化有关。raybet雷竞技最新例如,Bradshaw和Holzapfel[37]发现,在过去的24年里,北方猪笼草蚊(Wyeomyia smithii)的临界滞育光周期已经向南方转移。de Jong和Brakefield[38]发现,斑点瓢虫(Adalia bipunctata)的黑化倾向发生了变化,他们也将其与最近的气候变化联系起来。然而,在气候变化下,这种快速的进化反应可能是罕见的,因为适应气候变化需要高度的raybet雷竞技最新遗传多样性,而性状之间的共变异可能会减缓这一过程[39,40]。Gienapp等人[41]还警告说,其他声称气候变化下微进化证据的研究未能将遗传变化与表型可塑性分开。raybet雷竞技最新
既然我们已经研究了气候变化对昆虫种群的直接影响,我们将探讨气候变化的间接影响。raybet雷竞技最新这些间接的变化机制会导致昆虫与其食物或捕食者在空间和时间上的不匹配。这些不匹配最终会影响我们因昆虫而获得或失去的生态系统服务。
3.寄主植物对昆虫的介导作用
食草昆虫的生存高度依赖于与植物的相互作用。寄主植物通过物候和营养条件直接影响食草动物种群[42 46]。例如,Nzekwu和Akingbo-hungre[47]表明,利用不同类型的寄主植物对昆虫发育有显著影响。先前对蝴蝶的研究也表明,幼虫需要具有适当物候的持久寄主植物才能承受环境变化[48]。任何资源的变化都可能影响食草动物的动态和丰度。草食性昆虫与其食用植物之间的不匹配是由两类昆虫的不同反应引起的;例如,昆虫的早期出现和范围转移的速度可能比植物更快。这种不匹配会导致昆虫数量的下降。对于那些影响授粉等过程的物种来说,这可能会导致生态系统服务的减少。
3.1.昆虫与寄主植物间的时间不匹配
气候变化对昆虫的间接影响主要是由于寄主植物物候和raybet雷竞技最新品质的改变[50 53]。寄主植物质量下降导致死亡率增加,因为昆虫幼虫通过增加消耗量和发育时间来弥补营养价值的下降,这反过来又增加了它们对捕食者和其他环境压力的暴露[2,54]。此外,以短暂资源为食的物种更可能对与寄主植物[55]的异步生长敏感(例如,种群变异性增加)。例如,昆虫的卵在寄主植物的蓓蕾破裂之前孵化,它们可能会饿死,而那些需要嫩叶但孵化晚的昆虫将被迫吃防御更严密的叶子。
实验表明,温度升高对昆虫发育的影响大于对寄主植物的影响[57,58]。事实上,冬蛾(Operophtera brumata)之间已经出现了不同步现象。
和它的寄主植物有柄橡树(Quercus robur),因为O. brumata的卵孵化比Q. robur[59]的芽爆裂提前。然而,在这种特殊情况下,作者推测,O. brumata的高水平遗传变异可能会使Q. robur随着时间的推移迅速适应物候。大多数类群缺乏关于物种可塑性的信息;因此,很难预测这是否会是一个普遍的反应(参见Visser[60]关于适应气候变化的讨论)。raybet雷竞技最新
提供重要授粉服务的昆虫,例如蝴蝶,特别容易受到食物资源暂时不匹配的影响。平均而言,蝴蝶种类的发展速度比草本植物快[61]。例如,在英国,迁徙的红上将蝶(Vanessa atalanta)在过去几十年里提前了返程飞行,但它的寄主植物刺荨麻(Urtica dioica)并没有提前开花时间,造成了物候学上的不匹配[62]。在另一项研究中,McLaughlin等人将气候变化导致的降水变异性增加与海湾斑蝶(Euphydryas editha bayensis)两个种群的灭绝联系起来。raybet雷竞技最新这种可变性通过加速植物相对于幼虫发育的衰老而导致灭绝[28,48]。Memmott等人[63]的模拟得出结论,17 50%的传粉者(包括昆虫)将与它们的食物分离,与其他情况一样,较小的饮食宽度是与食物不同步的更大风险因素。稍后,我们将讨论其他对人类活动产生负面影响的物种如何从与捕食者的时间不匹配中受益。
3.2.昆虫与寄主植物的空间不匹配
在许多昆虫系统中,昆虫的传播能力大于它的寄主植物。专业食草动物的这种差异传播能力,如果它们仍然受到寄主植物的限制,可能会导致活动范围的收缩。许多昆虫有长距离移动的能力。例如,欧洲的一些蝴蝶在过去30年里向北移动了240公里。对于植物来说,最近的变化并没有很好的记录,但随着过去的气候变化,冰川后的范围扩张超过100公里/ 100年,中位数仅为20 - 40公里/世纪[64]。raybet雷竞技最新这并不奇怪,因为植物,尤其是大树,通常比昆虫的繁殖时间长,招募量低[65]。植物和昆虫之间的这些差异由于栖息地的破碎而进一步复杂化,由于其有限的传播能力,这可能对植物的迁移特别具有限制性[66 68]。
为了说明在气候变化下空间不匹配的潜在影响,我们分析了到达美国北部地理极限的特殊蝴蝶与其主要寄主植物的关系。raybet雷竞技最新共有来自15个亚科的74种蝴蝶,它们在美国北部的范围边界和利用单一的寄主物种。其中59种蝴蝶和寄主植物均有县级分布记录。
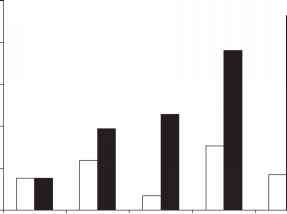
阴性0-50 50-100 100-500 500-1000 1000-2000
距离/公里
图1北美寄主蝴蝶最北部的地理分布与其幼虫寄主植物之间的空间距离分类基于观测蝴蝶和植被记录的县级质心位置的距离(ET SpatialTechniques, ArcGIS, Version 9.0)[71]。白色柱状表示每个类别内的总数,黑色柱状表示跨类别的相加总数。
阴性0-50 50-100 100-500 500-1000 1000-2000
距离/公里
图1北美寄主蝴蝶最北部的地理分布与其幼虫寄主植物之间的空间距离分类基于观测蝴蝶和植被记录的县级质心位置的距离(ET SpatialTechniques, ArcGIS, Version 9.0)[71]。白色柱状表示每个类别内的总数,黑色柱状表示跨类别的相加总数。
利用ET SpatialTechniques, ArcGIS, Version 9.0, ESRI计算了每个物种范围内最北端县[69]及其寄主植物[70]的地理形心。空间不连续距离由寄主植物的质心纬度减去蝴蝶种类的质心纬度计算(图1)。
在评估的59种特殊物种中,46%的地理范围分布在最北的100公里内,76%的地理范围分布在500公里内,93%的地理范围分布在1000公里内,7%的地理范围超出了宿主的范围边缘(图1)。其中,有10种(17%)在与宿主相同的位置达到了当前的活动范围极限。此外,9个物种(15%)的分布范围超出了它们当前的食用植物范围边缘(图1中为负值)。这可能是由于所使用的观测数据的规模引起的抽样误差,广泛的纵向分布导致多个北部范围前沿,由于偶尔迁移到下沉栖息地而导致的范围边界评估不准确,或物种的范围扩展未被记录。
假设寄主植物边际相对稳定,寄主对专业食草动物的偏好一致,蝴蝶和寄主活动范围边际的空间差异将决定蝴蝶活动范围在气候变化下能扩大到多远。raybet雷竞技最新在蝴蝶的边界与寄主之间的地理距离较小的地方,可能有一个较小的距离可供蝴蝶活动范围扩展。考虑到欧洲蝴蝶的活动范围变化程度(30 a以上240公里)[19],我们估计会上升
到2050年,北美59%的特殊蝴蝶将达到寄主的活动范围极限,假设寄主植物分布不变,无限扩散。例如,佛罗里达叶翼虫(Anaea floridalis)生活在距离其宿主分布边缘不到100公里的地方,它将在短短十多年内接近宿主边界,其他昆虫物种也将如此。北美蝴蝶动物群中高达4%的区域可能会经历缩小的地理范围,其中有限的食物资源分布与未来的气候变暖相互作用,从而阻止充分的气候跟踪。raybet雷竞技最新简单的风险评估,例如这种比较昆虫与寄主植物向极地分布边界的程序,可以帮助确定在气候变化下风险最大的物种。raybet雷竞技最新
既然我们已经探索了气候变化如何导致食草昆虫及其食物之间的时间raybet雷竞技最新和空间不匹配,我们转而研究气候变化如何使昆虫及其捕食者之间的相互作用脱钩,从而可能导致物种的增加和移动,从而通过爆发破坏栖息地。
4.捕食者对昆虫种群的影响
很少有研究考虑到气候变化对较高营养水平(即捕食者、拟寄生物和病原体)的影响及其对食草昆虫种raybet雷竞技最新群的后续影响。raybet雷竞技最新气候变化预计会通过独立改变每个营养类群的动态来影响捕食者和猎物之间的关系[71]。此外,研究发现,较高的营养级对气候变化更敏感,这可能是由于气候变化对较低营养级[7]的综合影响,以及较高营养级对非生物胁迫的更敏感[72 74]。raybet雷竞技最新顶端营养层的变化会引起营养级联。因此,捕食者的减少会破坏包括食草动物和初级生产者在内的较低营养水平之间的相互作用[75,76]。
捕食者,特别是拟寄生物,在控制昆虫数量方面发挥着重要作用[77 82]。在某些情况下,它们被发现会引起食草昆虫的周期性动态([82,83];但请见裁判。[80,84]为例外)。最近的研究森林昆虫已将种群暴发与昆虫生活史物候变化导致的气候变化联系起来[85 87]。raybet雷竞技最新然而,在气候变化下,较高的营养水平在引起昆虫爆发中的作用才刚刚开始得到解决[88]。raybet雷竞技最新我们回顾了迄今为止气候变化导致捕食者和猎物关系的时空不匹配及其对草食动物爆发的后续影响的观测和证据。raybet雷竞技最新
4.1.昆虫和捕食者之间的时间不匹配
草食动物的发育通常会随着气候条件的变化而迅速而显著地变化,这就导致了巨大的时态变化在他们的人口中。拟寄生物也可能受到气候变化及其对寄主动力学的影响。raybet雷竞技最新例如,Stireman等人[88]的一项研究使用了自然生态系统中饲养的毛虫的多年库存数据,将寄生频率与年际气候变化联系起来。raybet雷竞技最新他们发现,寄生蜂的寄生频率与降水的可变性呈负相关。这表明,在气候变化下,由于气候变率增加,导致拟寄生性草食动物相互作用不匹配,草食动物的爆发将会增加。raybet雷竞技最新
如果捕食者和它们的昆虫食草动物对气候变化的反应不同,就可能发生时间上的不匹配。raybet雷竞技最新寄主和拟寄生物可能有不同的热偏好或在极端温度下生存的不同能力,而对气候的不同反应可能会破坏同步[89]。raybet雷竞技最新例如,有证据表明拟寄生物的耐温性往往低于寄主[90,以及其中的参考文献]。模型预测,由于对气候的不同反应,捕食者和猎物之间物候不同步的增加可能导致种群动态的不稳定和系统的灭绝[91,92]。raybet雷竞技最新
然而,很少有研究从经验上证明了捕食者和猎物对气候变化的不同反应。raybet雷竞技最新Van Nouhuys和Lei[93]发现,局限于阴凉栖息地的浅色寄生虫Cotesia melitaearum发育缓慢,而其蝴蝶宿主Melitaea cinxia的深色幼虫则会寻找温暖的微栖息地,在那里阳光会提高体温,使个体发育更快。因此,在凉爽但阳光充足的春天,在大多数幼虫宿主已经化蛹之后,拟寄生物就出现了。在温暖的春季,寄主与寄生蜂保持同步,寄生率较高。在这项研究中,作者发现同步性对本地宿主种群大小没有影响,但同步性的中断可能对整体宿主元种群动态很重要。
有证据表明,在较高的营养水平上,对气候变化和其他形式的非生物胁迫越来越敏感[7,73,74,94]。这可能是由于气候变化对低营养水平的综合影响[7]。raybet雷竞技最新Menge和Sutherland[72]的群落调节理论预测,较高营养水平的生物对非生物胁迫更敏感,因为处于高营养水平的生物可能更大,流动性更强,使它们能够从次优区域分散出去。例如,人们发现,在灾难性事件后,营养水平较高的物种恢复速度较慢[95,96]。此外,Spiller和Schoener[94]发现,在1999年和2001年的两次飓风过后,十个加勒比小岛上的食肉动物减少,这可能解释了观察到的食草动物增加的原因。Preisser和Strong[74]还观察到,在与厄尔尼诺事件相关的2年降水低于平均水平后,加利福尼亚海岸的羽扇豆(lupine, Lupinus arboreus)大量死亡。羽扇豆的死亡是由于一种根食食草动物的爆发,加州小海蒲。加州小绥螨的种群由异缘线虫(nema-tode Heterorhabditis marelatus)[97]调节,其移动依赖于土壤湿度。在他们的研究中,Preisser和Strong通过实验将土壤水分含量提高到厄尔尼诺潮湿年份的水平(高于平均降水水平),发现水分水平的增加直接影响了掠食性线虫,但没有影响到扇豆平或鬼蛾。捕食者数量的增加抑制了加州小绥螨,间接保护了羽扇豆。 Climate change is expected to increase such extreme events [98].
4.2.昆虫和捕食者的空间不匹配
物种正在改变它们的活动范围以应对气候变化,但正如上面讨论的食草动物及其植物的运动一样,昆虫食草动物raybet雷竞技最新及其捕食者可以以不同的速度移动。依赖于与宿主密切联系的捕食者,如寄生虫和拟寄生物,预计尤其会受到宿主空间分布变化的影响。殖民寄主可能缺乏寄生虫或拟寄主,并且/或被感染的寄主可能不太适合,因此不太成功地在新的地点建立[99]。此外,新殖民地区的本土掠食者可能会从本土宿主转变过来;然而,一些本土捕食者可能需要在物候学、行为或生态方面进化出专门的物种,才能影响到外来物种。因此,捕食者物种复合体可能比本地范围内的捕食者较少,攻击率也较低[100 102]。在食草昆虫不断扩大的范围边缘,寄生蜂或其他捕食者的减少可能会通过摆脱捕食者的控制,给不断扩大的物种带来优势(例如,种群规模的增加)。
迄今为止,只有一项研究调查了气候变化下食草昆虫及其拟寄生物的不同传播率。raybet雷竞技最新Menendez等人[103]发现,由于气候变暖而向北迁移的棕阿格斯蝴蝶(Aricia agestis)在新殖民地区受到拟寄生物的攻击,死亡率较低。raybet雷竞技最新其他关于被引入新地区的入侵性昆虫或由于寄主植物的引入而扩大其活动范围的昆虫的研究也发现,在新殖民地区,拟寄生物的比例较低[100,104]。入侵物种的敌人在其入侵范围内的减少也已被记录为许多其他植物和动物物种(例如[105,106])。
Menendez等[103]发现在agestis的扩展范围和本地范围内,拟寄生蜂的丰富度没有差异;然而,他们确实发现了拟寄生物种组成的差异。在阿格斯提斯的新殖民范围内,大多数攻击它的物种都是已经出现在该地区的多面手,攻击一种本地蝴蝶Polyommatus icarus。其他研究发现,当昆虫宿主扩大它们的活动范围时,专性拟寄生物就消失了,而且在它们入侵的范围内攻击宿主的大多数物种都是从其他原生宿主转变而来的多面手[100,104,107]。
与从原生宿主转变而来的多面手寄生蜂相比,跟随宿主的专门性寄生蜂可能是更有效的捕食者。虽然有更多的多面手寄生蜂在其扩大的范围内攻击阿格斯蒂斯寄生蜂,但最丰富的寄生蜂被认为是历史上没有出现过的,因此很可能是一种与阿格斯蒂斯寄生蜂一起扩大其范围的专家[103]。其他关于入侵宿主的研究也发现了类似的结果——入侵范围内数量最多的捕食者是来自本土范围的专业捕食者[106]。
在扩大的范围内的通才物种可能对经历了范围扩大的新宿主来说是不那么有效的捕食者。例如,虽然在agestis的本地和扩展范围内,拟寄生蜂的物种丰富度相似,但与本地宿主P. icarus相比,agestis的攻击率较低[103]。最近入侵加州圣华金河谷的斑叶蝉(Erythroneura variabilis)也发现了类似的结果。与本地葡萄叶蝉E. elegantula[108]相比,E. variabilis受到共享拟寄生蜂的攻击率较低。因此,有证据表明,原生拟寄生物迁移到新宿主的速度很慢,因为它们在本地适应了原生宿主。
随着时间的推移,随着敌人追赶上扩大的宿主分布,在入侵范围内的捕食者和宿主的组合将有望变得类似于在原生范围内的组合。大多数研究发现,随着时间的推移,在宿主入侵或扩大的范围内,掠食者的丰富度会增加,但这导致了与宿主原生范围内不同的物种组成。Schonrogge等[104]发现,对于一些正在扩大活动范围的灰蜂,引入范围内的拟寄生蜂组合更多地由攻击现有灰蜂种类的拟寄生蜂组合决定,而不是由先前活动范围内的拟寄生蜂决定。
到目前为止,还没有研究将宿主范围转移后的捕食减少与经历了扩张的物种数量的增加联系起来。需要对正在其范围的不同部分进行范围扩张的物种的捕食者进行实验操作,以将扩大范围内拟寄生物的减少与种群动态的变化联系起来。入侵生物学文献提供了测试在扩大范围内减少敌人对宿主释放的框架[99,109,110]。然而,迄今为止还没有对昆虫扩大其活动范围以应对气候变化进行测试。raybet雷竞技最新由于许多昆虫都是重要的害虫,我们现在讨论气候变化如何有利于影响重要人类活动和生态系统服务的本地和非本地害虫,包括森林损raybet雷竞技最新失和碳固存减少。
5.raybet雷竞技最新气候变化和害虫
能够使一个物种对气候变化做出良好反应的特征正是害虫物种(本地和非本地)通常具有的特征。raybet雷竞技最新例如,害虫通常具有广泛的饮食范围、多voltine、快速生长、高流动性和/或表型可塑性[111 113]。雷竞技csgo由于许多害虫都具有这些特征,它们可能会对气候变化做出有利的反应,可能比那些特化和/或罕见的害虫反应更强烈。raybet雷竞技最新
例如,Chown等人[114]发现,与本土弹尾相比,入侵弹尾在应对变暖条件下干燥的能力方面更具表型可塑性。
害虫影响许多人类活动和生态系统服务。许多昆虫物种已经具有破坏性,在气候变化的影响下,有些物种可能会变得更加严重。raybet雷竞技最新害虫是北美森林中最重要的干扰因素,其影响面积几乎是火灾面积的50倍[18,115116],估计美国每年平均经济损失为21亿美元[117]。许多害虫是非本地的;在北美,多达40%的主要害虫种类是侵入性的[118],29种主要林业害虫中的62%是非本地的[117]。害虫,无论是本地的还是非本地的,不仅对经济有影响,而且对生态也有重大影响。例如,入侵凤仙花(Adelges piceae)造成了弗雷泽冷杉(Abies fraseri)残林的大量死亡,并威胁到其他依赖弗雷泽冷杉的本地物种[119]。
由于气候变化,许多害虫种类扩大了它们的活动范围。raybet雷竞技最新松树行蛾(T. pityocampa)是欧洲南部松树和其他针叶树的害虫,由于过去30年冬季气温上升,冬季存活率增加,它已经向北迁移了87公里,迁移到了更高的海拔地区。由于夏季和冬季气温升高和降水减少,山松甲虫(D. ponderosae)也向北移动到加拿大西部的高海拔地区[120]。除了扩大其活动范围外,这些害虫和其他害虫的爆发也更加频繁和持续时间更长[85 87]。虽然昆虫爆发的原因很复杂,但观察到的爆发严重程度的增加既与气候变化对昆虫生理的直接影响有关,也与通过寄主植物(例如,如果寄主植物更受干旱胁迫)和捕食者的变化所产生的间接影响有关(例如[85 88,121 123])。raybet雷竞技最新
最后,气候变化对森林害虫的影响可能会影响当前森raybet雷竞技最新林预算的平衡。D. ponder-osae引起的广泛树木死亡导致森林初级生产力降低,并因树木腐烂而增加碳排放。据预测,在未来20年里,加拿大不列颠哥伦比亚省的松林将成为净碳源,而不是净碳汇,这是由最近气候变化导致的D. ponderosae暴发的严重程度增加所介导的[86,87,124]。raybet雷竞技最新
6.结论
在这篇综述中,我们证明了气候变化对昆虫物种影响的方向和程度是多方面的。raybet雷竞技最新变化在空间和时间上都在发生,这些变化直接来自气候变化,间接来自与营养水平较低(即植物)和较高(即捕食者)的物种的相互作用。raybet雷竞技最新此外,营养级联可以发生,这样食草动物和它们的寄主植物之间的相互作用的变化会影响更高的营养水平,而捕食者和食草性猎物之间的变化会影响初级生产者。
此外,物种特征,如扩散能力、营养水平和专业化程度,可能是气候变化影响的良好预测指标。raybet雷竞技最新我们的预测是,未来的群落将由移动通才、繁殖时间快的物种、具有高度分散能力的物种或已被人类分散到全球各地的物种主导。这些物种中有许多已经是害虫,在气候变化下,新物种可能成为新的寄主植物或新地点的害虫。raybet雷竞技最新这些机会主义者可能会影响重要的人类活动,如林业和农业,并扩大到森林作为碳汇的作用。昆虫提供了重要的生态系统服务例如授粉或生物控制就不会有那么好的效果。
确认
本文得到了美国能源部科学办公室(生物与环境研究)的资助,资助编号为DE-FG02-05ER64023给JJH, DE-FG02-06ER64232给RLL。SLP主导的第2和第3部分;KMP主导的第4和第5段;图1由DJP和JDKD进行分析,JJH和RLL发起项目并编辑稿件。
参考文献
1.J.S.贝尔,G.J.马斯特斯,I.D.霍金森,C.奥马克,T.M.贝泽默,V.K.布朗,J.巴特菲尔德,A.巴斯,J.C.库尔森,J.法拉尔,j.e.古德,R.哈林顿,S.哈特利,T.H.琼斯,R. l .林德罗斯,M.C.普雷斯,I.西姆尼欧迪斯,ad .瓦特,J.B.惠特克,格洛布。《变化生物学》8 (2002)
2.T.M.贝泽默,T.H.琼斯,Oikos 82(1998) 212 222。
3.斯特灵,科内利森,格洛布。变更生物学。13(2007)1823 1842。
4.M.S. Warren, J.K. Hill, J.A. Thomas, J. Asher, R. Fox, B. Huntley, D.B. Roy, M.G. Telfer, S. Jeffcoate, P. Harding, G. Jeffcoate, S.G. Willis, J.N. Greatorex Davies, D. Moss, C.D. Thomas,自然41(2001)65 69。
5.N.R. Andrew, L. Hughes,生态学家。昆虫学报,29(2004)527 542。
6.c·a·多伊奇,j·j·图克斯伯里,r·b·休伊,k·s·谢尔登,c·k·加兰博,dc·哈克,p·r·马丁,纳特尔。学会科学。美国105(2008)6668 6672。
7.W. Voigt, J. Perner, A.J. Davis, T. Eggers, J. Schumacher, R. Bahrman, B. Fabian, W. Hein rich, G. Kohler, D. Lichter, R. Marstaller, F.W. Sander,生态学84(2003)2444 2453。
8.C. Parmesan, G. Yohe,自然421(2003)37 42。
9.T.L. Root, J.T. Price, K.R. Hall, S.H. Schneider, C. Rosenzweig, J.A. Pounds,自然421(2003)57 60。
10.O.戈多,J.J.桑兹,生态学家。昆虫学报,31(2006)261 268。
11.W.N. Ellis, J.H. Donner, J.H. Kuchlein, Ent。的误码率。72.中国科学(1997)66。
12.d·b·罗伊,T.H.斯帕克斯,格洛布。变更生物学。6(2000)407 416。
13.M.L.福斯特,上午夏皮罗,水珠。变更生物学。9(2003)1130 1135。
14.C. Stefanescu, J. Penuelas, I. Filella, Glob。变更生物学杂志。9(2003)1494 1506。
15.R.A.弗莱明,G.M.塔切尔,见:R.哈林顿,N.鹳(编),变化环境中的昆虫,学术出版社,伦敦,1995年,第505 508页。
16.R.哈林顿,S.J.克拉克,S.J.威尔汉姆,P.J.维里尔,C.H.丹农,M.胡勒,D.莫里斯,M.D.朗塞维尔,N.科库,格洛布。变更生物学。13(2007)1550 1564。
17.D.L.穆索林,格洛布。变更生物学。13(2007)1565 1585。
18.j。a。洛根,j。雷尼埃,j。a。鲍威尔,前线。生态。环境。1(2003)130 137。
19.C. Parmesan, N. Ryrholm, C. Stefanescu, J.K. Hill, C.D. Thomas, H. Descimon, B. Huntley, L. Kaila, J. Kullberg, T. Tammaru, J. Tennent, J. a . Thomas, M. Warren,自然299(1999)579 583。
20.L. Crozier,气raybet雷竞技最新候变化与物种范围边界:以大腹圆腹蛾(鳞翅目:大腹圆腹蛾科)北部范围界限为例,博士论文,华盛顿大学,华盛顿。
21.L. Crozier,生态学报135(2003)648 656。
22.J.K.希尔,Y.C.科林汉姆,C.D.托马斯,D.S.布莱克利,R.福克斯,D.莫斯,B.亨特利,生态学。let . 4(2001) 313 321。
23.C.D. Thomas, E.J. Bodsworth, R.J. Wilson, A.D. Simmons, z.g Davies, M. Musche, L. Con radt,自然411(2001)577 581。
24.R. Karban, S.Y. Strauss,生态学。昆虫学报29(2004)251。
25.A.T. Peterson, E. Martinez Meyer, C. Gonzalez Salazaar, P.W. Hall, Can。[J. Zool. 82(2004) 851 858]。
26.A. Battisti, M. Stastny, S. Netherer, C. Robinet, A. Schopf, A. Roques, S. Larsson,生态学。申请15(2005)2084 2096。
27.C. Parmesan, S. Gaines, L. Gonzalez, D.M. Kaufman, J. Kingsolver, A.T. Peterson, R. Sagarin, Oikos 108(2005) 58 75。
28.J.F. McLaughlin, J.J. Hellmann, C.L. Boggs, P.R. Ehrlich, Proc. Natl。学会科学。美国99(2002)6070 6074。
29.J.F. McLaughlin, J.J. Hellmann, C.L. Boggs, P.R. Ehrlich,生态学报132(2002)538 548。
30.j.k希尔,C.D.托马斯,R.福克斯,M.G.特尔弗,S.G.威利斯,J.阿瑟,B.亨特利,Proc. R. Soc。Lond。爵士。B, 269(2002) 2163 2171。
31.J.K.希尔,Y.C.科林汉姆,C.D.托马斯,D.S.布莱克利,R.福克斯,D.莫斯,B.亨特利,生态学。let . 4(2001) 313 321。
32.莱维坦,《Evol》。生态。第5(2003)号决议597 604。
33.P.A. Umina, A.R. Weeks, M.R. Kearney, S.W. McKechnie, A.A. Hoffmann,科学308(2005)691 693。
34.J.巴兰亚,J. m .奥勒,R.B.休伊,G.W.吉尔克里斯特,L.塞拉,科学313(2006)1773。
35.F. Rodriguez Trelles, M.A. Rodriguez, Evol。生态学报12(1998)829 838。
36.N.E. Rank, E.P. Dahlhoff,进化56(2002)2278 2289。
37.W.E. Bradshaw, C.M. Holzapfel, Proc. Natl。学会科学。美国98(2001)14509 14511。
38.德容,下午布雷克菲尔德,研究委员。Lond。爵士。B 265 (1998) 39
39.J.R. Etterson, R.G. Shaw,科学294(2001)151 154。
40.J.J. Hellmann, M. Pineda Krch, Biol。自然科学进展。137(2007)599 609。
41.贾纳普,杨晓华,杨晓华,杨晓华,杨晓华,杨晓华,《分子生物学》17(2008)167 178。
43.J.C. Schultz, P.J. Nothnagle, I.T. Baldwin, Am。J. Bot 69(1982) 753 759。
44.R.F.查普曼,见:R.F.查普曼,A. Joern(编),蚱蜢生物学,食物选择,威利,纽约,1990年,第36页72。
45.M.D. Hunter, P.W. Price,生态学73(1992)724 732。
46.P.W.普赖斯,见:M.D. Hunter, T. Ohgushi, P.W.普赖斯(编),资源分配对动植物相互作用的影响,学术出版社,圣地亚哥,加州,1992,第139 - 173页。
47.A.N. Nzekwu, A.E. Akingbohungbe, J.直翅目11目(2002)185 188。
48.j·j·赫尔曼,j·阿尼姆。生态学报71(2002)925 936。
49.N.J.桑德斯,D.M.戈登,生态学84(2003)1024 1031。
50.p·d·科利,克里姆。变更39(1998)455 472。
51.G.J.马斯特斯,V.K.布朗,I.P.克拉克,J.B.惠特克,J.A.霍利尔,Ecol。昆虫学报23(1998)45 52。
52.J.B.惠特克,欧洲。中国昆虫学报,39(1999):349。
53.S.E.哈特利,T.H.琼斯,Oikos 101(2003) 6 17。
54.C.V.约翰斯,A.休斯,格洛布。变化生物学。8(2002)142 152。
55.R.E. Forkner, R.J. Marquis, J.T. Lill, J.L.E. Corff, Ecol。昆虫学报,33(2008)276 285。
56.r·d·哈林顿,r·i·沃伍德,t·斯帕克斯,潮流生态。vol. 14(1999) 147 149。
57.R.C.杜瓦,瓦特,生态学89(1992)557 559。
58.j·k·希尔,I.D.·霍金森,生态学家。昆虫学报,20(1992)237 244。
59.m·范·阿施,p·h·范·蒂恩德伦,l·j·m·霍勒曼,m·维瑟,格洛布。变更生物学。13(2007)1596 1604。
60.维瑟,教授。Lond。爵士。B 275(2008) 649 659。
61.C.帕尔马干酪,格洛布。变更生物学13(2007)1860 1872。
62.T.H.斯帕克斯,d。b。罗伊,r。l。h。丹尼斯,格洛布。变更生物学。11(2005)507 514。
63.J. Memmott, P.G. frenzy, N.M. Waser, M.V. Price, Ecol。第10号(2007)710 717。
64.M.B.戴维斯,R.G.肖,科学292(2001)673 679。
65.杜灵格,丁博克,格拉伯尔,生态学杂志。92(2004)472。
66.j·k·希尔,c·d·托马斯,b·亨特利Lond。爵士。B 266(1999) 1197 1206。
67.L.R.艾弗森,A.普拉萨德,M.W.施瓦茨,生态。115(1999) 77 93。
68.O. Honnay, K. Verheyen, J. Butaye, H. Jacquemyn, B. Bossuyt, M. Hermy,生态学家。Lett. 5(2002) 525 530。
69.P.A. Opler, H. Pavulaan, R.E. Stanford, M. Pogue,北美蝴蝶和飞蛾,Bozeman, MT, NBII山地草原信息节点,2006,可从http://www.butterfliesandmoths.org/。
70.美国农业部,NRCS,植物数据库,国家植物数据中心,巴吞鲁日,洛杉矶,2008,可从http://plants.usda.gov.
71.W.H.范德普特,P.C.德鲁特,T.M.哈维,m.w assen, V. Wolters,《基础应用》。生态学报5(2004)487 494。
72.曼格,j.p·萨瑟兰,美国Nat 130(1987) 730 757。
73.L. Cagnolo, S.I. Molina, G.R. Valladares,生物潜水员。保护11(2002)407 420。
74.E.L. Preisser, D.R. Strong, Am。Nat 163(2004) 754 762。
75.M.L.佩斯,J.J.科尔,S.R.卡朋特,J.F.基切尔,趋势生态。vol. 14(1999) 483 488。
76.G.A.波利斯,A.L.W.西尔斯,G.R.赫塞尔,D.R.斯特朗,J.马龙,趋势生态。vol. 15(2000) 473 475。
77.M.P.哈塞尔,J.拉托,R.M.梅,J.阿尼姆。生态学报58(1989)883 892。
78.康奈尔,霍金斯,美国Nat 145(1995) 563 593。
79.霍金斯,康奈尔,霍赫伯格,生态学78(1997)2145 2152。
80.亨特医生,瓦莱医生,格雷德威尔医生,纳特教授。学会科学。美国94(1997)9176 9181。
81.P. Turchin, A.D. Taylor, J.D. Reeve,科学285(1999)1068 1071。
82.P. Turchin, S.N. Wood, S.P. Ellner, B.E. Kendall, W.W. Murdoch, A. Fischlin, J. Casas, E. McCauley, C.J. Briggs,生态学84(2003)1207 1214。
84.J.利布霍尔德,J.埃尔金顿,D.威廉姆斯,R.M.穆兹卡,波普。生态学报42(2000)257 266。
85.D.W.威廉姆斯,上午的信息环境。昆虫学报,24 (1995)19
86.M.P. Ayers, M.J. Lombardo, Sci。总环境。262(2000)263 286。
87.W.J.A.沃尔尼,R.A.弗莱明,农业公司。Ecosyst。环境科学。82(2000)283 294。
88.J.O.斯蒂尔曼三世,L.A.戴尔,D.H.简森,M.S.辛格,J.T.利尔,R.J.马奎斯,R.E.里克莱夫斯,G.L.金特里,W.哈尔瓦克斯,P.D.科利,J.A.巴隆,H.F.格林尼,H.康纳斯,P.巴博萨,H.C.莫莱斯,R.迪尼兹,纳特尔。学会科学。美国102(2005)17384 17387。
89.T.汉斯,J.范·巴伦,P.弗农,G.鲍文,安努。中国生物工程学报,2007,27(2)。
90.嘉雅港,田田雅,安。Entomol。Soc。Am. 62(1969) 1303 1306。
91.F. Herard, M.A. Keller, W.J. Lewis, J.H. Tumlinson, J. Chem。生态14(1988)1583 1596。
92.h·c·j·戈弗雷,m·p·哈塞尔,r·d·霍尔特,j·阿尼姆。Ecol. 63 (1994) 1
93.范努赫斯,雷,阿尼姆。生态学报5(2004)526 535。
94.41. d.a Spiller, T.W. Schoener,生态学88(2007)37。
95.R.D. Holt,见:G.A. Polis, K.O.酿酒师(编),食物网:模式和动态的集成,查普曼和霍尔,纽约,1996年,第313 326页。
96.d.a Spiller, J.B. Losos, T.W. Schoener,科学281(1998)695 697。
97.D.R.斯特朗,A.V.惠普尔,A.L.蔡尔德,B.丹尼斯,生态学80(1999)2750 2761。
98.IPCC, S. Solomon,秦丹,M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor, H.L. Miller(编),气候变化2007:自然raybet雷竞技最新科学基础。第一工作组对政府间气候变化专门委员会第四次评估报告的贡献,剑桥大学出版社,英国剑桥和纽约,2007年。raybet雷竞技最新
99.托钦法医,米歇尔法医,前线。生态。环境科学。2(2004)183 190。
One hundred.康奈尔,霍金斯,美国Nat 141(1993) 847 865。
101.J. b . f . Geervliet, M. s . w . Verdel, J. Schaub, H. Snellen, M. Dicke, L.E.M. Vet,生态学报124(2000)55 63。
102.M.沃斯,L.E.M.兽医,Evol。生态。第6(2004)1021 1035号决议。
103.r.m enendez, A. Gonzalez Megias, O.T. Lewis, M.R. Shaw, C.D. Thomas, Ecol。昆虫学报,33(2008)413 421。
104.施罗尼格,P.沃克,M.J.克劳利,普罗克。Lond。B生物学265(1998)1643 1650。
105.C.E. Mitchell, A.G. Power,自然421(2003)625 627。
106.M.E. Torchin, K.D. Lafferty, A.P. Dobson,自然421(2003)628 630。
107.K. Schonrogge, G.N. Stone, M.J. Crawley, Oikos 77(1996) 507 518。
108.W.H.塞特尔,L.T.威尔逊,生态学71(1990)1461 1470。
109.K.谢伊,P.切森,趋势生态。vol. 17(2003) 170 176。
110.R.I. Colautti, A. Ricciardi, L.A. Grigorovich, H.J. MacIsaac, Ecol。Lett. 7(2004) 721 733。
雷竞技csgo111.D.B. Simberloff,见:J.A. Drake (Ed.),生物入侵:一个视角,威利,纽约,2005年,第61 - 75页。
112.J.H.劳顿,K.C.布朗,Proc. R. Soc。Lond。爵士。B 314(1986) 607 617。
113.N.L.沃德,G.J.马斯特斯,格洛布。变更生物学。13(2007)1605 1615。
114.S.L. Chown, S. Slabber, M. McGeoch, C. Janion, H.P. Leinaas, Proc. R. Soc。Lond。爵士。B 274(2007) 2531 2537。
115.V.H. Dale, L.A. Joyce, S. McNulty, R.P. nelson, M.P. Ayres, M.D. Flannigan, P.J. Hanson, L.C. Irland, A.E. Lugo, C.J. Peterson, D. Simberloff, F.J. Swanson, B.J. Stocks, B.M. Wotton,生物科学51(2001)723 733。
116.G.M. Lovett, C.D. Canham, M.A. Arthur, K.C. Weathers, R.D. Fizhugh,生物科学56(2006)395 405。
117.D. Pimental,见:D. Pimental(编),生物入侵。外来植物、动物和微生物物种的经济和环境成本,CRC出版社,博卡拉顿,佛罗里达,2002年,151 155页。
118.P. Niemela, M.J. Mattson,生物科学46(1996)741 753。
119.点利布霍尔德,W.L.麦克唐纳,D.贝里达尔,V.C.马斯特罗。科学。科学通报,30 (1995)
120.A.L. Carroll, S.W. Taylor, J. Regniere, L. Safranyik,见:T.L. Shore, J.E. Brooks, J.E. Stone(编),挑战与解决方案,报告编号BC X 399,加拿大自然资源,加拿大森林服务,维多利亚,2004年,第223 232页。
121.E.E. Berg, J.D. Henry, C.L. Fastie, ad . De Volder, S.M. Matsuoka, For。生态。管理。227(2006)219 232。
122.K.F. Raffa, B.H. Aukema, B.J. Bentz, A.L. Carroll, J.A. Hicke, M.G. Turner, W.H. Romme,生物科学58(2008)501 517。
123.L.C.斯蒂格,陈嘉生,张志强,D. Frank, N.C. Stenseth,纳特尔。学会科学。美国104(2008)16188 16193。
124.W.A. Kurz, C.C. Dymond, G. Stinson, G.J. Rampley, E.T. Neilson, A.L. Carroll, T. Ebata, L. Safranyik,自然452(2008)987 990。
继续阅读:海洋生物——作为气候和全球变化指标的远洋和浮游生态系统raybet雷竞技最新
这篇文章有用吗?