气候与全球变化的潮间带指标raybet雷竞技最新
Nova Mieszkowska
英国海洋生物协会,实验室,城堡山,普利茅斯PL1 2PB,英国
1.简介
2.raybet雷竞技最新气候变化与生物地理学
2.1.利用长期数据集探测气候变化raybet雷竞技最新
2.2.潮间带生物群的反应
2.3.极端天气事件
2.4.的相互作用
3.机制
3.1.耐热性
3.2.繁殖和招募
4.全球变化的额外影响
4.1.海洋酸化
4.2.沿海发展
4.3.raybet雷竞技最新气候变化与非本地物种
5.致谢及参考文献
1.介绍
岩石潮间带横跨的地区的海岸线从最高垂直水平达到高潮期间大潮(伴随着海浪飞溅)到低水位泉水暴露在空气中的最低水平。岩石潮间带栖息着各种各样的类群,包括藻类、软体动物、棘皮动物、刺胞动物和甲壳动物。由于岩石海岸容易接近,潮间带物种在整个19世纪和20世纪被业余博物学家[1,2]和专业研究人员作为生态和生物学理论发展的模型系统进行了广泛的研究[3 15]。
潮间带无脊椎动物和大型藻类是海洋进化起源的变温动物,但由于它们每天都要出现和浸泡在海水中,它们必须同时适应海洋和陆地的环境。因此,他们
raybet雷竞技最新气候变化:观测到的对地球的影响
版权所有©2009 Elsevier B.V.保留一切形式的复制权利。
提供对水生和陆地气候环境变化的影响的独特见解。潮汐昼夜循环以及海气温度的季节性波动意味着潮间带生物会受到极端温度的影响,导致体温经常出现超过30°C的波动[16]。干燥[17]、水流和波浪力[18]、盐度的快速波动[19 21]、氧气可用性[22,23]和营养水平[24]等额外的压力因素意味着生物体通常生活在接近其生理耐受极限的地方[25 32]。
海洋变温物种通常比陆地物种对环境变化的反应更快:典型的短寿命和成年和幼年阶段的固定或不动的性质阻止了逃离不断变化的环境制度。大多数潮间带物种的幼虫阶段是浮游的,因此也提供了环境变化对海洋的影响远洋区.因此,分布和丰度的变化很可能是由生物体对环境变化的直接反应所驱动的。潮间带无脊椎动物和海洋大型藻类来自较低的营养水平,因此预计它们比营养水平较高的物种对当地条件的变化做出更快的反应,通常在食物链向上到三级和顶端捕食者的级联效应中表现出第一个反应[33,35]。关键结构或功能物种丰度的变化会改变整个岩石群落的组成和动态[5,36,37],而这些环境条件的微小变化会导致群落结构和功能的重大改变[38,39]。综上所述,岩质潮间带生态系统可能是最敏感的自然系统之一,也是对气候变化最早做出反应的生态系统之一[40,41]。raybet雷竞技最新
潮间带物种的地理范围基本上是一维的,因为它们占据了低潮和高潮之间的狭长海岸线[42]。结合这些物种生活的高度紧张和波动的环境,岩石潮间带因此是研究气候变化影响的理想模型系统。raybet雷竞技最新岩石潮间带生态系统遍布全球,因此便于对全球环境变化的相对影响进行时空比较。对环境变化的反应可以分为两类;近似的生态响应,依赖于非生物因素与生物水平过程、种群动态和群落结构之间的关系,并通过生理、形态和行为的变化直接影响个体在不同生命阶段的表现。这些影响可扩大到种群水平的响应,这还可能受到气候驱动的水文过程变化的影响,这些变化影响到远洋幼虫生命阶段的扩散和招募。raybet雷竞技最新所有这些都会导致分布、生物多样性、生产力和微进化过程的改变。
2.raybet雷竞技最新气候变化与生物地理学
生物地理学研究由Tournefort[44]在18世纪首次提出,20世纪初开展的工作[24,45 51]被用作今天生态气候变化研究的基础。raybet雷竞技最新主要的海洋生物地理区主要是根据感兴趣的分类群的生物地理分布界限的聚类来确定的。海水温度被认为最终决定了海洋物种的生物地理范围(见[24,52])。低纬度和高纬度生物地理界限分别与8月和2月的海面等温线有关,包括海藻[53 58]、卷脚动物[59]和软体动物[60,61]等不同分类类群的动植物都有相同的界限。然而,物种分布与气候之间的关系并不简单。raybet雷竞技最新生物地理学研究常常因为环境参数的变化而变得复杂,这阻碍了人们对因果关系的理解。
适宜的栖息地存在于许多海洋无脊椎动物的分布范围之外[62 65],但目前不适宜的环境条件阻止了它们的殖民,因此认为其范围受到气候的限制。raybet雷竞技最新这一原则被称为一个物种的“气候包络线”,是当今使用的许多生物raybet雷竞技最新气候模型的基础[66,67],但参见参考文献。(68、69)。当环境条件改变到一个物种的生理耐受范围内时,随着生物能够在合适的栖息地开拓新的地点,预测范围将会扩大。然而,在实践中,范围边缘可能位于这个基本生态位“包络线”内的一段距离。物种之间、生物与环境因素之间的相互作用,以及缺乏适宜栖息地、适宜栖息地空间的分散和连通性差等局部影响,都为每个物种确定了已实现的生态位。因此,必须测量气候变率对动植物分布及其相互作用的影响,以便了解并最终预测海洋生态系统的变化。
2.1.利用长期数据集探测气候变化raybet雷竞技最新
世界上存在一些最广泛的空间和时间数据集,用于东北大西洋沿岸潮间带无脊椎动物和大型藻类的分布和丰度。fisher - piette[70 - 73]在20世纪30年代、40年代和50年代沿着法国、西班牙、葡萄牙和北非的大西洋海岸线进行了密集和广泛的调查。Crisp和Southward在20世纪50年代对英国和爱尔兰的海岸线进行了类似的调查([74,75],Southward和Crisp,未发表数据)。这些数据集在气候变化监测方面特别有价值,因为它们提供了广泛的基线,可用于测量过去70年气候变暖和变冷期间潮间带物种分布和丰度的raybet雷竞技最新变化速度和程度[76]。英国海岸也有关于藤壶、tro-chids和帽贝的丰度和种群结构的时间序列数据,分别可追溯到20世纪50年代、70年代和80年代[74,75,77 79]。
海洋生物多样性和气候变化项目“MarClim”由英国海洋raybet雷竞技最新生物协会于2001年建立,旨在评估和预测气候变化对英国和爱尔兰岩石潮间带生物多样性的影响。它结合了历史数据和对400多个岩石海岸的当代重新调查(图1),提供了与近期气候变化相关的潮间带物种数量、种群结构和地理分布变化的证据[64]。raybet雷竞技最新MarClim调查方案与20世纪50年代的原始调查[77,75]所使用的相同,以绘制50多种无脊椎动物和大型藻类的分布和范围界限,包括冷水和温水来源。此外,还收集了藤壶、帽贝和球虫等关键物种的丰度和种群动态的定量数据。这些调查是在从范围边缘到更接近分布中心的地点进行的。这些联合数据集已被用于跟踪暖水和冷水物种在海岸上的丰度和相对优势的变化,在整个20世纪,它们共同存在以应对波动的气候条件。
2.2.潮间带生物群的反应
由于全球环境变化,地理范围边缘的收缩和扩张导致物种从群落中消失或被引入群落。这种变化最初记录在一个物种地理范围的外围,那里的生物通常已经经历了接近其热极限的温度[26]。然而,在一个物种的地理范围内也可能存在局部或区域异质性,这可以从远离固着无脊椎动物分布极限的环境热点[31,40]或冷点[79]得到证明。这些变化反过来又影响物种相互作用的结果,如竞争、促进和捕食,最终改变群落结构和海洋生态系统过程[41,43,80 82,85]。
自20世纪80年代中期气候开始迅速变暖以来,英国已经发生了大范围潮间带类群分布范围的变化。raybet雷竞技最新自20世纪以前的记录以来[64、65、83],温暖水域的扬子腹足类(如Osilinus lineatus和Gibbula umbilalis),藤壶类(包括Chthamalus montagui, C. stellatus和Perforatus (Balanus) Perforatus)和棕色巨藻Bifurcaria bifurcata)的北部和东部范围边缘已经扩展到85至180公里之间,并且速度非常快
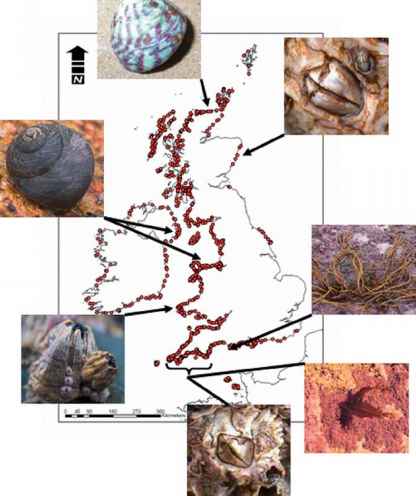
图1 20世纪50年代和21世纪初收集的英国岩石海岸的调查数据。从左下顺时针方向,是温水物种,它们已经扩展到了北部的边界;穿孔虫,线腹鼠,脐带Gibbula, Chthamalus montagui, Bifurcaria bifurcata。在其南部范围边缘表现出收缩的冷水种:咸水藻,半圆藻。
图1 20世纪50年代和21世纪初收集的英国岩石海岸的调查数据。从左下顺时针方向,是温水物种,它们已经扩展到了北部的边界;穿孔虫,线腹鼠,脐带Gibbula, Chthamalus montagui, Bifurcaria bifurcata。在其南部范围边缘表现出收缩的冷水种:咸水藻,半圆藻。
每十年可行驶50公里。这一速度远远快于陆地物种每十年6.1公里的平均移动速度[84],并且与英国沿海水域浮游生物的北移相一致(见本卷第12章)。帽贝Patella rustica最近在葡萄牙北部沿海水域上升流减弱期间弥补了其分布的历史空白[79]。造成这种现象的原因可能有几个,包括海面温度的升高、比斯开湾南部上升流强度的减弱以及西伊比利亚大陆架海流强度的增加,所有这些都是由全球气候驱动的。raybet雷竞技最新
在过去的十年里,北欧的大面积地区已经消失,斯堪的纳维亚半岛、黑尔戈兰和布列塔尼南部海岸的种群数量大幅减少。然而,在英国海岸线上这种巨藻的数量没有记录到这样的下降[64],这表明其原因可能不是气候因素,而是潜在的疾病相关因素。
在地中海,从大西洋流入的温暖海水和热带物种的繁殖体,以及从红海通过苏伊士运河迁移的物种较少,以及人类对外来物种的引入正在改变生态系统结构,并对营养网产生潜在影响[86]。海洋洞穴还受到温暖水域mysid增加和特有冷水同类严重减少的影响[87]。
在过去的几十年里,冷水起源的物种包括藤壶(Semibalanus bala-noides)[88]、龟壳帽贝(Tectura testudinalis)和棕色巨藻(Alaria esculenta)[66,89]在英国和欧洲的南部范围限制和丰度下降。
在20世纪50年代、60年代[90,91]和21世纪初[92]期间,对葡萄牙沿海大型藻类分布的调查已经确定了-120种显著物种,这些物种在这些时期之间的范围边缘位置发生了重大变化。自1941年以来,暖水物种的分布移动与年平均近海海温(SST)之间存在显著相关性[93]。冷水起源的物种表现为南向和北移,但作为一个群体来看没有明显变化。这种不同的反应可能部分是由于本研究对这些物种进行了分组,其中一些是非本地物种,而另一些则被广泛认为在整个欧洲具有世界性分布,而不是温暖或寒冷的亲缘关系[76]。
蓝贻贝Mytilus edulis在1977年至1994年之间从挪威大陆向北延伸了500公里,到达斯瓦尔巴特群岛的熊岛[94],并在2002年至2004年异常温暖的大西洋海水涌入后,于1000年首次在北极斯瓦尔巴特岛被发现[95,96]。这种重新出现代表了分布范围向极地移动的巨大变化-1000公里,可能是由于上层幼虫在温暖地区向北移动造成的水流.然而,目前尚不清楚这些种群是否可持续,或者当前的气候是否仍然太冷,使该物种无法在如此高纬度地区繁殖和生存。raybet雷竞技最新
2.2.3.美国
对美国大西洋和太平洋海岸岩石海岸的研究可以追溯到20世纪初,但大多仅限于最近几十年,这源于人们越来越意识到需要广泛的时空覆盖数据集来跟踪和预测全球环境变化的影响[97,98]。地理和海洋学对美国太平洋沿岸潮间带群落结构都有很大的影响。在温暖的年份,一些来自东太平洋加利福尼亚生物地理省的物种已经将它们的活动范围向北扩展到高纬度地区[99,100]。在加利福尼亚州蒙特利湾的一个固定地点,从20世纪30年代早期到90年代,温水腹足类、帽贝和花藻类的丰度增加而冷水丰度减少,推断出了其他的生物地理变化[38,102]。这些共生物种相对优势的改变改变了组合组成,使其成为一个更典型的温水群落。20世纪70年代末/ 80年代初至21世纪初,南部新腹足动物Kelletia kelletii在东北太平洋的加利福尼亚地区的生物地理范围向北移动[103,104]。从19世纪30年代至今的化石记录和调查表明,这是第一次有记录的延伸超出概念点,并且与20世纪后期海温的强烈变暖相吻合[104]。自20世纪70年代以来,由于夏季海水温度上升到足以促进其成功繁殖的程度,美国东海岸缅因湾出现了一种温水绿藻——易碎藻(Codium fragile)[105]。
2.2.4.南半球
由于澳大利亚和新西兰长期与其他温度地区地理隔离,因此它们都有非常多的地方病[106,107]。在这些地区,物种数量和地理范围的减少可能导致物种在全球范围内灭绝。在澳大利亚,已经启动了新的研究项目,以跟踪岩质潮间带物种的生物地理迁移速率[108],但该地区的数据仍然很少。随着沿海水温的升高,东海岸温带地区已经发现了热带岩池鱼类[109]。由于海水温度上升的直接影响,塔斯马尼亚岛从低潮间带到浅潮下形成密集地带的温带海带物种一直在持续减少[110111]。雷竞技csgo自20世纪60年代以来,来自大陆的温水长刺海胆(Centrostephanus rodgersii)的大量传播,以及温水海胆(Janus edwardsii)和鲍鱼(Haliotis rubra)的大量增加,加剧了这种下降[108]。西部岩龙虾,Parulirus cygnus是澳大利亚渔业中最重要的单一物种[113]。渔业登陆量与吕因流的强度密切相关,吕因流推动幼虫的跨大陆架运输,从而提高种群的生产力[113]。吕温流强度与ENSO事件高度相关,在El Niño年减弱。自20世纪70年代以来,El Niño事件变得更加常见[114],导致龙虾渔业规模出现更频繁的萧条。
新西兰几乎没有覆盖整个海岸线的潮间带物种的定量数据,尽管个别地点存在广泛的时间序列([115]和Ballantine,未发表的数据)。对新西兰的定量调查始于2008年多岩石的海岸线建立一个基线,以衡量未来气候引起的物种分布变化和生态系统层面的反应[116]。raybet雷竞技最新实验表明,在新西兰海岸,贻贝Perna canaliculus对温暖空气温度的耐受性低于共生的Mytilus spp.[117],并且在移植到更高的潮间带海拔时,其生长和繁殖能力下降,这表明气候变化的潜在有害影响。raybet雷竞技最新这个物种通常居住在低海岸和潮湿的,中间海岸的阴影区域,因此可能不会受到像实验操作条件下那样严重的暴露。
2.2.5.南非
非洲大陆几乎没raybet雷竞技最新有发表与气候相关的研究。暖水岩潮间带种帽贝(Patella longicosta, P. oculus)和螺贝(Oxystele variegata)的活动范围扩大和种群水平变化。在南非的冷水帽贝P. granatina中观察到招募失败是对南部异常温暖的表面温度的响应目前本格拉(在南非附近)在1982/1983年[118]可能与1982/1983年的强地震有关厄尔尼诺事件在太平洋[119]。虽然有大量的证据表明物种扩大了它们的北部活动范围,但很少有证据表明它们缩小了南部活动范围。这一问题部分源于对许多潮间带物种的南部生物极限过去和现在的位置缺乏了解,以及从南部极限种群收集的数据缺乏,这些种群往往位于非洲或南美沿海水域。
2.3.极端天气事件
的极端寒冷由于暴露在零度以下的温度下,英国1962/1963年冬季大大缩短了许多潮间带物种的北部活动范围。威尔士北部的社区受到的影响尤其严重,因为在凌晨气温最低的时候出现了低潮。威尔士和英格兰南部的人口受影响较小,因为正午左右出现低潮,这些地区的北部界限被重新划定[120]。威尔士北部的O. lineatus的北部范围边缘并没有从极端造成的退缩中恢复多少寒冷的冬天从1962年到1963年直到20世纪80年代。在这中间的20年里,其分布范围在海岸线周围重新扩大了-100公里,多年龄、繁殖期的种群已经在以前的限制范围内15公里内建立起来[121]。
在当前全球环境快速变化的时期,热浪事件正变得越来越普遍,由于暴露在高温下,热浪事件还导致了包括太平洋海星Pisaster ochra-ceus (Harley,个人通讯)、贻贝M. edulis和大型藻类(Mieszkowska,个人观察)在内的潮间带物种的灾难性死亡。有趣的是,P. ochraceus的生存和觅食行为似乎没有受到长期的、不太严重的空气温度上升的负面影响[122],这表明只要强烈的热冲击不频繁发生,P. ochraceus将在很长一段时间内应对全球变暖。尽管极端天气事件可以暂时改变潮间带物种的活动范围,但似乎是长期气候趋势决定了生物地理界限和大规模分布变化。
2.4.的相互作用
越来越多的物种从温暖气候地区正开始取代那些在时间区域具有较冷气候亲和力的,导致局部组合的组成改变[76,101,1raybet雷竞技最新02,123,124]。这些局部尺度的变化还将通过改变范围边缘种群内灭绝与殖民事件的比率,促进物种向极地的传播。在英国和北欧,相对于南部温水藤壶(Chthamalus species)和帽贝(P. depressa),北部冷水藤壶S. balanoides和帽贝Patella vulgata的数量也有所下降[76,125,126]。使用英国长期藤壶时间序列建立和测试的模型表明,这一速度正在增加,预计在2060年至2080年期间,英格兰西南部的S. balanoides将完全被Chthamalus spp.所取代。raybet雷竞技最新气候变化也在改变促进性的相互作用。冷水帽贝,P. vulgata,优先把自己的家设在大藻类vesiculosus的阴影下。为了模拟气候变暖的影响,实验中从海岸移除泡囊F. vesiculosus,观察到P. vulgata的显著死亡率,幸存的个体重新迁移了他们的家车。raybet雷竞技最新相比之下,温水同族P. depressa的死亡率不会增加,也不会改变其位置[72,127]。
3.机制
虽然相关生物地理学研究可用于获得物种发生的概率图并产生可检验的假设,但它们不能提供关于因果关系的信息[84,128]。为了准确预测未来物种分布的生物地理变化的速度和程度,需要更好地了解驱动这些变化的生物学机制。物理、生态、进化和生理因素作用于繁殖、出生、扩散、招募和死亡过程,这些因素都参与了物种范围的形成[25,129 131],在研究环境变化的影响时也必须考虑到这些因素。从细胞和分子到生物组织的有机体和种群水平,已经发现raybet雷竞技最新了对气候变暖的机械反应。
3.1.耐热性
由于热休克蛋白(hsp)的产生增加,大多数植物和动物都能够承受一定程度的热应激,这种蛋白在升高或降低温度下逆转细胞蛋白质的展开,并从细胞中移除严重受损的蛋白质[132]。热休克蛋白表达增加的地理趋势已经在潮间带物种中显示出来,如Nucella canaliculata,一种温带潮间带海螺,出现在东北太平洋的岩石海岸线上。在南部种群中hsp表达的增加表明个体在南部范围边缘比在范围中心更受压力[133]。紫色海肠(Strongylocentrotus purpuratus)在响应温度升高的热sp诱导的遗传调控方面也表现出地理差异,这反映了沿生物地理分布的不同位置所经历的热应激水平[134]。因此,HSPS可能是监测潮间带物种受气候影响程度的敏感工具。raybet雷竞技最新
3.2.繁殖和招募
人们早就知道,海洋和空气温度的变化也会影响海洋物种的生理性能和繁殖成功[14,45,137 141]。自20世纪80年代中期以来,整个英国和法国北部的南部扬轮鸟的数量都有同步增加的记录。这种数量的增加与20世纪80年代中期以来地区气候变暖有关,这是由于每年招募成功的频率增加[65]。raybet雷竞技最新这些变化背后的机制是,南部棘足腹足类Osilinus lineatus和Gibbula脐腹足类为了应对温和的冬季和温暖的春季而提前开始每年的繁殖周期,再加上新定居的新物种(通常是对环境压力最敏感的生命阶段)暴露在温和、较短的冬季时的生存能力增加[121]。在英国西南部,南方帽贝的年度繁殖周期也开始得更早,持续时间更长。相比之下,只有不到20%的北帽贝达到了可以产卵的性腺发育阶段[135]。招募也会受到海洋特征的影响,海洋特征可以控制远洋幼虫阶段的扩散。对美国太平洋海岸的大规模调查表明,海表温度与招募之间存在密切的相关性,沿生物地理范围的关键石藤壶和贻贝物种的招募率存在很大的种内差异[98],这表明气候变暖可能会改变招募成功的可能性。raybet雷竞技最新
4.全球变化的额外影响
4.1.海洋酸化
在21世纪下半叶,海洋酸化预计将对藤壶、帽贝和顶壳等岩岸物种产生影响。海洋变暖和海洋pH值降低的潜在协同效应尚不清楚,也可能因生物在物种纬度分布中的位置而异。
4.2.沿海发展
全球环境变化的其他方面包括海平面上升和极端天气事件[142]。世界各地许多低洼的海岸线地区正面临着更大的洪水风险。这种风险导致了沿海防御设施的激增,以保护财产,农业用地以及公路和铁路等基础设施。当沿着陆地-海洋界面的大片区域建造多个结构时,局部防御可以扩大到整个海岸线。这种情况发生在地中海[143,144]、爱尔兰海北部海岸、英吉利海峡东部和北海。这些发展可能会造成严重的影响,导致沉积栖息地的丧失,取而代之的是生物群落贫瘠的人造岩石海岸[144]。这种大规模的海岸改造也会影响生物地理过程。最近,英吉利海峡东部岩石海岸物种的活动范围扩大,可能是繁殖成功率提高和提供人工栖息地(海上防御设施、码头、海滨码头)作为跨越不利栖息地的垫脚石共同作用的结果[145]。
4.3.raybet雷竞技最新气候变化与非本地物种
由于海运的增加,非本地物种的引入在全球范围内不断增加,水产养殖、海水养殖、水族贸易及进口活鱼饵。压舱水、船体污垢和密集养殖已被确定为将入侵性非本地物种引入海洋环境的高风险载体。也有证据表明,这种引入更有可能发生在一个更温暖的世界[146]。为英国水产养殖和海水养殖业务进口贝类物种(例如,太平洋牡蛎长牡蛎,地中海蓝贻贝
M. galloprovinicialis)促进了这些物种在英国、欧洲和美国的自然生态系统中的传播。近岸海水温度现在足够温暖,允许这些引进物种成功繁殖,导致它们在种植地点以外的地区大举殖民,往往不利于本地同类,因为它们在竞争中被非本地物种击败。一旦外来物种在自然环境中扎根,就几乎没有什么障碍可以防止进一步传播。来自不同生物地理领域的非本土物种的引入可以被认为是全球环境变化的一个方面,有可能导致生物的全球同质化,从而降低生物多样性。
5.结论
潮间带无脊椎动物和藻类已经对全球气候变暖做出了反应,生物地理分布从温暖的低纬度地区向较冷的两极转移。raybet雷竞技最新总的来说,北方物种的后退速度不如南方物种的前进速度快。变化的速度和程度也是物种特有的,导致群落组成的改变,对生态系统结构和功能产生连锁反应,并可能导致温带热带地区生物多样性的下降。这些变化最终是由对温度的生理反应驱动的,此外还有来自物种相互作用、海洋过程、海岸带开发和栖息地可用性的间接影响。目前仍缺乏具有长时间和宽空间覆盖的数据,这阻碍了探测沿海生态系统的环境驱动变化,特别是在南半球。至关重要的是,研究应侧重于将维护这些有价值的数据集与开展实验研究相结合,以确定所观测到的响应的机制,以便我们能够更好地预测我们快速变化的环境对潮间带系统的未来影响。
确认
作者希望感谢K.理查德森的评论和建议,这有助于改进本章;MarClim项目www.mba.ac.uk marclim和MarClim基金财团;以及B.T. Helmuth、B.R. Broitman和E.S. Poloczanska在出版方面的协助。
参考文献
1.P.H. Gosse, Tenby:海边度假,John van Voorst, Paternoster Row,伦敦,1856年,第1397页。
2.C.金斯利,格劳克斯;或者,《海岸奇观》,麦克米伦和公司,剑桥,1856年,第1 168页。
8.T. Carefoot,太平洋海岸,J.J.道格拉斯,温哥华(1977)1 208。
9.R.T. Paine, J.M. Levine, Ecol。51(1981) 145 178。
10.J.R.刘易斯,《岩石海岸的生态学》,英国大学出版社,伦敦,1964年,第1323页。
11.p.k.代顿,生态学家。单41(1971)351 389。
12.门格生态文学士单,46(1976)355 393。
13.A.J. Southward,自然175(1955)1124 1125。
14.A.J.索思沃德,J. 3 .比尔。Assoc。UK 36 (1957) 323 334
16.B.T.赫尔穆特,M.W.丹尼,Limnol。Oceanogr.48(2003) 1338 1345。
18.J.A.基茨,M.W.丹尼,比尔。牛。8(2005)114 119。
19.J.达文波特,H.马卡利斯特,J. Exp. marbiol。生态学报76(1996)985 1002。
20.李荣华,李荣华,李荣华,中国生物医学工程学报(自然科学版)。
21.L.E.伯内特,美国Zool. 37(1997) 633 640。
22.R.F.服务,科学305(2004)1099。
23.E.P.达尔霍夫,b.a Menge, 3月生态学家。掠夺。爵士144(1996)97 107。
24.l·w·哈钦斯生态学院单17(3)(1947)325 335。
25.R.M. MacArthur,《地理生态学:物种分布的模式》,Harper & Rowe,纽约。269页。
26.J.R.刘易斯,水生物学142(1986)1 13。
27.J.H.布朗,G.C.史蒂文斯,D.M.考夫曼,安努。启生态。系统27(1996)597 623。
29.J.H.斯蒂尔曼,G.N.塞米罗,杰布。199 (1996) 1845
30.A.A.霍夫曼,P.A.帕森斯,《极端环境变化与进化》,剑桥大学出版社,1997年,第235页。
31.B.赫尔穆特,C.D.哈利,下午Halpin, M. O'Donnell, G.E. Hofmann, C.A. Blanchette,科学298(2002)1015 1017。
32.j·h·斯蒂尔曼,integrg公司。生物学报42(2002)790 796。
33.M.H. Carr, J.E. Neigel, J.A. Estes, S. Andelman, R.R. Warner, J.L. Largier, Ecol。应用13,S90 S107。
34.P.E.史密斯,坎。j .鱼。Aquat。科学42(1985)69 82。
35.S. Jenouvrier, C. Barbraud, H. Weimerskirch, JAE 72(2003) 576 587。
36.j·r·刘易斯,3·比尔。安。Rev. 14(1976) 371 390。
37.S.J.霍金斯,R.G.哈特诺尔,J. Exp. Mar. Biol。生态学报62(1982)271 283。
38.j.p Barry, C.H. Baxter, R.D. Sagarin, S.E. Gilman Science 267(1995) 672 674。
39.j·r·刘易斯,r·s·鲍曼,m·a·肯德尔,p·威廉森,内斯。22 .海洋法第16号决议(1982年)
40.B.赫尔穆特,N.米什科夫斯卡,P.摩尔,S.J.霍金斯,安努。启生态。另一个星球。系统37(2006)373 404。
41.M.D. Bertness, G.H. Leonard, J.M. Levine, J.F. Bruno,生态学报120(1999)446 450。
42.R.D. Sagarin, S.D. Gaines, J.生物地理学29(2002)985 997。
43.C.D.G.哈利,A.R.休斯,K. Hultgren, B.G. Miner, C.J.B. Sorte, C.S. Thornber, L.F. Rodriguez, L. Tomanek, S.L. Williams, Ecol。Lett. 9(2006) 228 241。
44.j·p·德·图尔内福特,《黎凡特航行关系》(1717年)。巴黎。288页。
46.j·h·奥顿,j·3·比尔。Assoc。英国2(1920)299 366。
47.W.A.塞切尔,海藻地理分布中的温度区间(1920)。科学187 190。
48.凯恩,基金会植物地理,哈珀兄弟,纽约,伦敦,1944年。
49.l·w·哈钦斯生态学院一夫一妻17(1947)325 335。
50.E.V.伍尔夫,《历史植物地理学导论》(由e.b里森登从俄语翻译而来),马萨诸塞州沃尔瑟姆,1950年。
51.H.G. Andrewartha, L.C. Birch,《动物的分布和数量》,芝加哥大学出版社,芝加哥,1954年。
52.E.C. Pielou,《生物地理学》,Wiley Interscience, Chichester, 1979。
53.点布里曼,Helgoland海洋研究42(1988)199 241。
55.K. Luhning,海藻的环境,生物地理和生态生理学(1990)John Wiley & Sons,伦敦,161。
56.G.M.沃斯科博尼科夫,上午布里曼,C. van den Hoek, V.N. Makarov, E.V. Shoshina, Bot。3月39日(1996)341 346。
57.f·j·莫雷纳,上午布里曼,J. Phycol. 33(1997) 330 343。
58.奥法尼迪斯,A.M.布里曼,J. Phycol. 35(1999) 919 930。
59.A.J. Southward,《自然》165(1950)408。
60.D.R. Franz, A.S. Merrill, Malacologia 19(1980) 209 225。
61.G.J.维尔梅吉,进化36(1982)561 580。
62.J.R.刘易斯,《岩石海岸的生态学》,英国大学出版社,伦敦,1964年。
63.M.A.肯德尔,J.R.刘易斯,水生物学142(1986)15 22。
64.N.米什科沃斯卡,R.利珀,P.摩尔,M.A.肯德尔,M.T.巴罗斯,D.李尔,E.波洛赞斯卡,K.希斯科克,P. s .莫斯拉,R. c .汤普森,R.J.赫伯特,D.拉佛利,J.巴克斯特,A.J.索斯沃德,S.J.霍金斯,J. 3 .比尔。Assoc。英国临时出版,20(2005)1 55。
65.N. Mieszkowska, M.A. Kendall, S.J. Hawkins, R. Leaper, P. Williamson, N.J. Hardman Mountford, A.J. Southward,水生物学555(2006)241 251。
66.r·g·皮尔逊,t·p·道森,格洛布。生态。生物地质12(2003)361 371。
67.G.R. Walther, E. Post, P.康威,A. Menzel, C. Parmesan, T.J.C. Beebee, T.J.C. Fromentin, O.H. Guldberg, F. Bairlein,自然416(2002)389 395。
68.A.J. Davies, L.S. Jenkinson, J.H. Lawton, B. Shorrocks, S. Wood,自然391(1998)783 786。
69.r·w·布鲁克,r·w·特拉维斯,e·j·克拉克,c·戴森,j·希尔。生物学报。245(2007)59 65。
70.E.菲舍尔·皮耶特,J.海奇。巴黎79 (1935)
71.E.菲舍尔·皮耶特,J.林。Soc。Zool. 40(1936) 181 272。
72.E.费舍尔·皮耶特,安。Insitut。海洋。摩纳哥31(1955)37 124。
73.D.J. Crisp, E. Fischer Piette, Ann。Insitut。海洋。摩纳哥36(1959)276 381。
74.A.J. Southward, D.J. Crisp, Proc. R. Ir。学术57 (1954)1
75.D.J. Crisp, A.J. Southward, J. mari Biol。Assoc。英国37(1958)157 208。
76.a。j。索思沃德,s。j。霍金斯,m。t。巴罗斯,j。瑟姆。生物学报20(1995)127 155。
77.M.A.肯德尔,J.R.刘易斯,水生物学142(1986)15 22。
78.M.A.肯德尔,J.莫尔。种马53(1987)213 222。
79.F.P.利马,N.奎罗斯,P.A.里贝罗,S.J.霍金斯,A.M.生物地理学家桑托斯,33(2006)812 822。
80.B.赫尔穆特,J.G.金索沃,E.卡灵顿,安。物理学报67(2005)177 201。
81.C. Parmesan, S. Gaines, L. Gonzales, D.M. Kaufman, J. Kingsolver, A.T. Peterson, R. Sagarin, Oikos 108(2005) 58 75。
82.P.摩尔,S.J.霍金斯,R.C.汤普森,3 .生态学。掠夺。第334号爵士(2007)11 19
83.R.J.H.赫伯特,S.J.霍金斯,M.希德,A.J.索斯沃德,J. 3 .比尔。Assoc。UK 83(2003) 73 82。
84.C. Parmesan, G. Yohe,自然421(2003)37 42。
85.I.巴契、C.温克、K.比肖夫、C.M.布赫霍尔茨、B.H.巴克、A.埃格特、P.费尔普菲尔、D.哈涅特、S.雅各布森、R.坎儿兹、U.卡斯滕、M.莫利斯、M. y .罗莱达、H.舒伯特、R.舒曼、K.瓦伦丁、F.温伯格、J.威斯。海带属植物的研究进展,生态学杂志,43(1)(2008)86,doi:10.1080/09670260701711376。
86.C.N. Bianchi,水生生物学580(2007)7 21。
87.P. Chevaldonne, C. Lejeusne,生态学。第6号(2003)371 379号。
88.D.S. Wethey, s.a Woodin,水生物学606(2008)139 151。
89.T. Vance, MRes论文,普利茅斯大学,2005。
90.F. Ardre,波特酒。生物学报,10(1970)1423。
91.F. Ardre,公牛。科学研究中心比亚里茨8(1971)359 574。
92.F.P.利马,P.A.里贝罗,N.奎罗斯,S.J.霍金斯,A.M.桑托斯,水珠。改变生物学13(12)
(2007) 2592 2604。
93.R.T.莱莫斯,H.O.皮雷,实习生。J. Clim. 24(2004) 511 524。
94.J.M. Weslawski, M. Zajaczkowski, J. Wiktor, M. Szymelfenig,极地生物学,18(1997)45 52。
95.萨维森,诺。Geogr。56 . Tidsskr. 56 (2002)
96.J. Berge, G. Johnsen, F. Nilsen, B. Gulliksen, D. Slagstad, 3 . Ecol。掠夺。爵士303(2005)167 175。
97.c·a·布兰切特,c·m·米纳,p·t·雷蒙迪,d·洛舍,k·e·k·黑迪,b·r·布伊特曼,j·生物地理学家。(2008)出版。
98.b·r·布罗伊特曼,c·a·布兰切特,b·a·门格,j·卢布琴科,p·a·雷蒙第,c·克伦茨,m·福利,d·洛舍,S.D.盖恩斯,eccol . monogr。78(3)(2008) 403 421。
99.R.S.伯顿,进化52(1998)734 745。
One hundred.M.E.海尔伯格,D.P.鲍尔奇,K.罗伊,科学292(2001)1707 1710。
101.j.p Barry, C.H. Baxter, R.D. Sagarin, S.E. Gilman,科学267(1995)672 674。
102.R.D. Sagarin, J.P. Barry, S.E. Gilman, C.H. Baxter, Ecol。学报。69(1999)465 490。
103.T.J. Herrlinger, Veliger 24(1981) 78。
104.D. Zacherl, S.D. Gaines, S.I. Lonhart, J.生物地理学。30(2003)913 924。
105.l·g·哈里斯,m·c·泰瑞尔,比尔。《入侵3》(2001)9
106.G.C.B. Poore,澳大利亚海洋环境状况报告:海洋环境技术附件:1由Leon P. Zann编译,昆士兰汤斯维尔大堡礁海洋公园管理局,海洋救援2000计划,堪培拉环境、体育和领土部,1995 ISBN 0 642 17399 0。
107.土池荣,西村勇,中国地质大学学报(自然科学版)
108.E.S.波洛赞斯卡,R.C.鲍科克,A.巴特勒,A.J.霍布迪,O.霍格·古尔伯格,T.J.昆兹,R.马泰尔,D.A.米尔顿,T.A.奥凯,A.J.理查德森,海洋学家。3月的杂志。安。启45
(2008) 407 478。
109.S.P.格里菲思,东海岸。大陆架科学58(2003)173 186。
110.K.S. Edyvane,《澳大利亚环境最终报告》,第一产业、水和环境部,霍巴特,2003年。
雷竞技csgo111.G.J.埃德加,C.R.萨姆森,N.S.巴雷特,康瑟夫。生物学报19(2005)1294 1300。
112.C. Johnson, S. Ling, J. Ross, S. Shepherd, K. Miller,澳大利亚塔斯马尼亚水产养殖和渔业研究所,FRDC项目编号2001/004,2005。
113.N.卡普提,C.丘布,A.皮尔斯,3 .弗莱肖。第52(2001)号决议1167 1174。
114.k·e·特伦伯斯,公牛。点。满足。第78(12)(1997)2771 2777。
115.J.E.莫顿,V.J.查普曼,利地区的岩石海岸生态,北奥克兰,奥克兰大学出版社,奥克兰,1968年,44页。
116.米什科夫斯卡,伦德奎斯特,弗莱许。审议决议。
117.L.E. Petes, b.a Menge, G.D. Murphy, J. Exp. Mar. Biol。生态学杂志(2007)83 91。
118.G.M.布兰奇,s.j.科学80(1984)61 65。
119.R.T.巴伯,F.P.查维斯,科学222(1983)1203 2110。
121.N.米什科夫斯卡,S.J.霍金斯,M.T.巴罗斯,M.A.肯德尔,J. 3 .比尔。Assoc。英国87(2007)537 545。
122.桑福德,J. Exp. 3 . Biol。生态学杂志(2002)199 218。
123.麦高恩,蔡尔顿,康伟,《中国鱼类营养报告》,37(1996)。
124.S.J. Holbrook, R.J. Schmitt, J.S.J. Stevens, Ecol。应用7(1997)。第1299 - 1310页。
125.A.J.索思沃德,J. 3 .比尔。Assoc。英国71(1991)495 513。
126.S.J. Hawkins, P. Moore, M. Burrows, E. Poloczanska, N. Mieszkowska, S.R. Jenkins, R.C. Thompson, M. Genner, A.J. Southward,水生物学,气候研究,37 (2/raybet雷竞技最新3),(2008)pp. 123 133。
127.P.摩尔,R.C.汤普森,S.J.霍金斯,J. Exp. 3 . Biol。生态学报344 (2007)170 180
128.M.J. Fortin, T.H. Keitt, B.A. Maurer, M.L.锥形,D.M. Kaufman, T.M. Blackburn, Oikos 108(2005) 7 17。
129.R.N.卡特,S.D.普林斯,自然293(1981)644 645。
130.D.M.洛奇,下午卡里耶娃,J.G.金索沃,R.B.休伊(编),生物相互作用与全球变化,辛诺,桑德兰,马萨诸塞州,1993年,第367 387页。
131.j·j·列侬,j·r·g·特纳,d·康奈尔,Oikos 78(1997) 486 502。
132.S.林奎斯特,安。修订生物化学55(1986)1151 1191。
133.C.J.B. Sorte, G.E. Hofmann, 3 . Ecol。掠夺。第274号(2004)263 268。
134.C.J.奥索维茨,G.E.霍夫曼,J. Exp. 3 . Biol。生态学报327(2005)134 143。
135.P. Moore,博士论文,普利茅斯大学,2005。
136.V.L. Loosanoff,科学102(1945)124 125。
137.R.A. Boolootian,见:R.A. Boolootian(编),棘皮动物生理学,威利,纽约,1966,第561 614页。
138.B. Cocanour, K. Allen,生物化学。物理学。20(1967)327 331。
139.R.E.斯蒂芬斯,Biol。牛。142(1972)132 134。
140.P.W.弗兰克,3月生物学31(1975)181 192。
141.B.T.赫尔穆特,G.E.霍夫曼,比尔。公牛201(2001)371 381。
142.IPCC,《raybet雷竞技最新气候变化2007:科学基础》,IPCC 2007 1996。
143.L.艾罗尔迪,M.阿比亚蒂,M.W.贝克,S.J.霍金斯,P.R.琼森,D.马丁,P.S.莫舍拉,A. Sundelöf, R.C.汤普森,P.阿伯格,海岸。工程52(2005)1073 1087。
144.点Moschella, M. Abbiati, P.¿Aberg, L. Airoldi, J. Anderson, J.M. Bacchiocchi, F. Bulleri, G.E. Dinesen, M. Frost, E. Gacia, L. Granhag, P.R. jonsonn, M.P. Satta, A. Sundelöf, R.C. Thompson, S.J. Hawkins, Coast。工程52(2005)10531071。
145.S.J. Hawkins, N. Mieszkowska, P. Moschella,未发表资料。
146.J.J. Stachowicz, J.R. Terwin, R.B. Whitlatch, R.W. Osman, PNAS 99(2002) 15497 15500。
继续阅读:植物生态学作为气候和全球变化的指示器raybet雷竞技最新
这篇文章有用吗?
推荐项目


读者的问题
-
肯尼斯•亨特2个月前
- 回复