水生禾草状的
灯芯草(Juncus spp.)、莎草(Carex spp.)和芦苇(Phragmites, Scirpus, Bolboschoenus)是在溪流和湖泊边缘发现的最常见的物种群。灯芯草(灯芯草属)也普遍存在于排水不良的牧场。尽管它们在潮湿的土壤中常见,灯心草(灯心草属)通常不耐受长时间的缺氧,迄今为止测试的该属所有物种的根状茎在放置在厌氧培养箱中几天后死亡。越冬灯芯草的一个显著特征是


图8.14 Scheuchzer棉草(Eriophorum scheuchzeri) -北极和高山湿地莎草,从阿尔卑斯山到北极间断分布。在这张照片中,它生长在格陵兰岛东北部(北纬73°)Mesters Vig的一个沼泽中。北极种群的E. scheuchzeri已被发现具有耐缺氧性,这一特性在非北极种群的E. angustifolium和E. vaginatum中未被发现(Crawford et al., 1994)。这些相对耐缺氧和耐洪水的植物在北极冬季的许多个月里,根茎都被包裹在冰中,因此必须忍受长时间的缺氧,直到初夏冰融化时,它们才突然重新暴露在氧气中。
图8.14 Scheuchzer棉草(Eriophorum scheuchzeri) -北极和高山湿地莎草,从阿尔卑斯山到北极间断分布。在这张照片中,它生长在格陵兰岛东北部(北纬73°)Mesters Vig的一个沼泽中。北极种群的E. scheuchzeri已被发现具有耐缺氧性,这一特性在非北极种群的E. angustifolium和E. vaginatum中未被发现(Crawford et al., 1994)。这些相对耐缺氧和耐洪水的植物在北极冬季的许多个月里,根茎都被包裹在冰中,因此必须忍受长时间的缺氧,直到初夏冰融化时,它们才突然重新暴露在氧气中。
整个冬天都有绿色的芽基。因此,在一些冬季光合作用的帮助下,根与芽之间的通气可以在它们的生存中发挥重要作用。相比之下,灯心草和普通芦苇(Schoenoplectus spp.和
芦苇(Phragmites australis),栖息在经常遭受长时间和深冬季洪水的缺氧泥浆中的地区,其根状茎能够在长时间的缺氧中生存(Braendle & Crawford, 1999)。
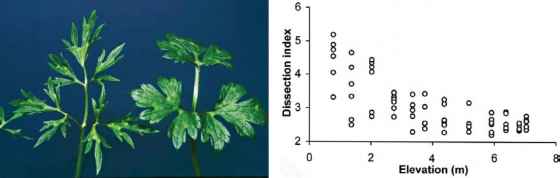
图8.15匍匐金凤花(Ranunculus repens)叶面解剖的种群差异。(左)在爱尔兰克莱尔郡霍克希尔特洛海拔最低的地方(左)和海拔最高的地方(右)种植了一年的R. repens叶子。(图片由Dr . Lynn提供)(右)来自田间和栽培植物的叶片解剖指数与透平海拔的关系。(经Lynn & Waldren授权转载,2003年出版)
图8.15匍匐金凤花(Ranunculus repens)叶面解剖的种群差异。(左)在爱尔兰克莱尔郡霍克希尔特洛海拔最低的地方(左)和海拔最高的地方(右)种植了一年的R. repens叶子。(图片由Dr . Lynn提供)(右)来自田间和栽培植物的叶片解剖指数与透平海拔的关系。(经Lynn & Waldren授权转载,2003年出版)

图8.16一个高寒沼泽,广泛分布着高寒克鲁姆霍尔兹松(Pinus mugo ssp.)的耐洪水形态。pumilio)。这个沼泽位于捷克共和国最高的山峰斯尼兹卡的山顶之下,海拔1602米(5300英尺),位于Krkonose山脉。的山脉也被称为巨大的山脉(德语,Riesengebirge)。捷克的名字很古老,出现在一个民族的名字中托勒密Corconti。山脉从西北延伸到东南,形成了波兰和捷克共和国的边界。(图片来自Tomas Kucera博士)
图8.16一个高寒沼泽,广泛分布着高寒克鲁姆霍尔兹松(Pinus mugo ssp.)的耐洪水形态。pumilio)。这个沼泽位于捷克共和国最高的山峰斯尼兹卡的山顶之下,海拔1602米(5300英尺),位于Krkonose山脉。山脉也被称为巨人山脉(德语,Riesengebirge)。捷克的名字很古老,出现在托勒密的一个民族的名字中,称为Corconti。山脉从西北延伸到东南,形成了波兰和捷克共和国的边界。(图片来自Tomas Kucera博士)

图8.17芦苇金丝雀草(Phalaris arundinacea),一种半水生沼泽草,具有根茎,在越冬时能耐受长期缺氧。在生长季节,如果洪水很浅,或者像这里流动的河流一样通气良好,这个物种能够使淹没在水中的器官通气。
图8.17芦苇金丝雀草(Phalaris arundinacea),一种半水生沼泽草,具有根茎,在越冬时能耐受长期缺氧。在生长季节,如果洪水很浅,或者像这里流动的河流一样通气良好,这个物种能够使淹没在水中的器官通气。
因此,耐缺氧的物种可以进一步分为两组,即那些可以在完全缺氧的情况下延长其枝条的物种,以及那些可以忍受长时间的缺氧,但等待地下水位下降和土壤通气恢复后才在春季恢复生长的物种(见表8.1)。在这两种极端的行为类型之间,有许多与强制缺氧有关的中间产物。在冬季耐缺氧的植物在初夏会变得易受缺氧的影响,因为它们的碳水化合物储备已经减少,以支持春季生长(见Phalaris arundinacea;图8.17)。这些变化说明了微妙的生态平衡,在物候学中,物种在洪水风险期间保持长时间的缺氧耐受,而更机会主义的策略则有利于在春季迅速恢复生长,即使由于后期洪水有周期性生长抑制的风险。
8.7.1 Glyceria maxima vs Filipéndula ulmaria
这种情况可以从草甸女王(Filipendula ulmaria)和芦苇甜草(Glyceria maxima;图8.18)。在苏格兰Tentsmuir国家自然保护区的一系列沙丘和松弛群落中,连续绘制这两种物种在24年内的分布图,这些群落的排水系统发生了变化,能够记录已经发生的变化的生态后果(图8.19,8.20)。洪水状况的变化在春季尤其明显,那时洪水较浅,消失得比过去快。对植被的影响是G. maxima将其领土扩展到以前由F. ulmaria主导的地区。大甘油菊对缺氧的耐受性不如大甘油菊,但像上面提到的许多容光属植物一样,在冬季保持绿色的基叶,如果淹水不深,将在春天在大甘油菊之前恢复生长。后者依赖于它的耐缺氧生存,直到水位下降到土壤表面以下,它才延长新一季的嫩芽。因此,当淹水较浅时,由于更早熟的物候,较不耐受的物种具有竞争优势。当淹水时间更长,深度维持在10厘米或以上时,大甘油无法与耐缺氧的F. ulmaria竞争(Studer-Ehrensberger et al, 1993;无花果,8.21 - -8.22)。
在湿地栖息地越冬需要有能力在长时间的淹没中生存,这对一些物种来说可能还包括长时间的缺氧。从上面的讨论可以明显看出,不仅植物淹水部分的耐缺氧能力对生存很重要,而且它与植物整体的关系也很重要。有或没有可通风的茎,或基生枝中残留的一些活动,甚至是树皮中的光合作用活动湿地树例如普通桤木,都可以减轻冬季缺氧的风险。潜在竞争者的存在也会影响洪水是否会为耐洪水物种创造一个边缘环境。
8.7.2甜旗(菖蒲)
甜菖蒲(Acorus calamus)是一种不可育(n = 36,三倍体)的新生植物(图8.23),可能是从一种1574年属于维也纳著名草药学家查尔斯·德·埃克卢斯(拉丁语为Clusius, 1525-1609;见Schröter, 1908)。然而,尽管明显缺乏群体杂合性
-
, an example of a semi-aquatic grass with numerous vegetative shoots and stout rhizomes spreading over wide areas in marshes and banks of slow running rivers.")
- 图8.18芦苇草(Glyceria maxima),半水生草的一个例子,具有大量的营养芽和粗壮的根茎,在沼泽和缓慢流动的河岸上广泛分布。
, an example of a semi-aquatic grass with numerous vegetative shoots and stout rhizomes spreading over wide areas in marshes and banks of slow running rivers.")

图8.19法夫岛Tentsmuir沙丘和松弛系统鸟瞰图。泥沙的快速堆积使这条海岸线迅速向海推进,形成了一系列沙丘和松弛群落,这些群落显示出受地形、营养物质和环境控制的明显边界水的供应.黄色方框表示图8.20所示样条的位置。(剑桥大学航空图片集版权所有)
图8.19法夫岛Tentsmuir沙丘和松弛系统鸟瞰图。沙粒的快速堆积使这条海岸线迅速向海推进,形成了一系列沙丘和松弛群落,这些群落受地形、养分和水供应的控制,呈现出明显的边界。黄色方框表示图8.20所示样条的位置。(剑桥大学航空图片集版权所有)
是欧洲异养湖泊边缘具有竞争性和侵略性的入侵者。这种入侵的成功似乎部分源于在这个物种的根茎中发现的极端耐缺氧。根状茎全年有高碳水化合物储备,足以维持几个月的乙醇发酵。此外,ATP的产量大大高于马铃薯(1.55-2.33 imol ATP g_1fr.wt)。h_1)和腺苷酸池以及能量电荷水平在长时间缺氧期间保持稳定和高水平(Sieber & brendle, 1991)。这些数字表明ATP的生产和消耗之间的平衡状态,加上代谢率足够高,允许组织在缺氧下延长生存能力。此外,根状茎的多孔性及其在湖泥表面的位置允许过量的乙醇扩散到湖水中,并避免了上面讨论的缺氧后转化为乙醛的危险(Studer & Braendle, 1984)。糖酵解和糖酵解相关酶的信使rna在冬季人工缺氧和自然环境下被诱导厌氧(Bucher et al., 1996)。在实验室和野外缺氧条件下形成的蛋白质已被证明是高度活跃的。此外,与非耐氧物种相比,除了直接参与厌氧代谢的蛋白质外,许多额外的蛋白质也在野外和实验室缺氧条件下合成,这清楚地表明,与马铃薯相比,缺氧对代谢的干扰较小,也不太可能导致细胞功能障碍(Armstrong et al., 1994)。 In addition, rhizomes of A. calamus are able to store and detoxify nitrogen that has been taken up as ammonium by the roots by transfer into alanine. The main nitrogen storage compound, however, in winter rhizomes is the nitrogen-rich amino acid arginine (Haldemann & Braendle, 1986, 1988). Arginine is readily converted into transport amino acids in spring when growth starts (Weber & Braendle, 1994). Nitrogen回收与其他沼泽植物相比,持续摄取有利于该物种的生长发育和蛋白质合成。类似的策略用于厌氧土壤中形成的硫化物的解毒和利用。它以谷胱甘肽的形式储存在根状茎中(Weber & brendle, 1996)。谷胱甘肽被用作硫源,但除了生育酚和酚类物质外,也可用于抗氧化防御机制(Larson, 1988)。该物种在耐缺氧方面最突出的策略可能是膜脂在缺氧下的稳定性及其在组织再充气时对过氧化损伤的保护(Henzi & Brandle, 1993)。在70天的缺氧后,脂质的饱和水平只有很小的变化,主要的变化是脂肪酸饱和度从18:3亚麻酸转变为18:2亚油酸。此外,组织中的游离脂肪酸很少,几乎没有膜分解的证据。
相比之下,在耐缺氧的普通梅花(Schoenoplectus lacustris)中,根状茎在缺氧35天后才开始出现一些损伤迹象。此外,在根状茎中只产生少量的过氧化产物丙二醛和乙烷。这个物种的膜很明显
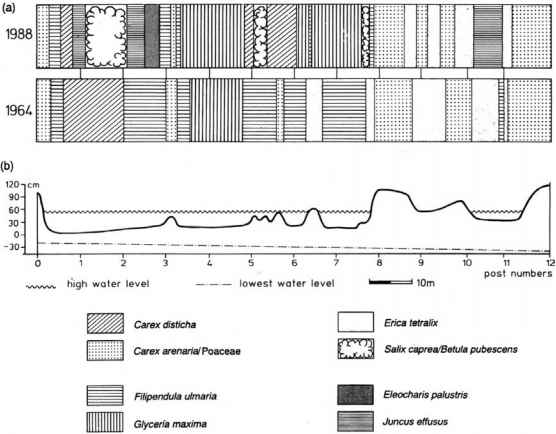
图8.20 Tentsmuir国家自然保护区(a) 1964年至(b) 1988年24年间记录的沙丘和松弛样带植被与地形的关系。大致位置见图8.19。1964年至1988年之间的主要变化是由于新的排水沟降低了洪水水平,耐缺氧程度较低的大甘油(Glyceria maxima)取代了耐缺氧程度较高的Filipendula ulmaria。(经Studer-Ehrensberger等人授权转载,1993年。)
图8.20 Tentsmuir国家自然保护区(a) 1964年至(b) 1988年24年间记录的沙丘和松弛样带植被与地形的关系。大致位置见图8.19。1964年至1988年之间的主要变化是由于新的排水沟降低了洪水水平,耐缺氧程度较低的大甘油(Glyceria maxima)取代了耐缺氧程度较高的Filipendula ulmaria。(经Studer-Ehrensberger等人授权转载,1993年。)
能很好地适应长时间的缺氧。缺氧条件下脂质代谢与蛋白质代谢不同,脂质被保存下来,而蛋白质可以重新合成。考虑到脂质的合成需要去饱和酶和分子氧,这种区别并不出乎意料。因此,菖笋在抵御缺氧的危险方面表现得特别好,无论是在对缺氧的内抗性方面,还是在对土壤溶液中外部产生的厌氧产物的抗性方面,它都可以被认为比芦苇和互花米草等耐缺氧的物种更适应缺氧,因为这些物种的高硫化物浓度可能会造成损害(见下文)。
8.7.3芦草(Glyceria maxima)
在两栖芦草(Glyceria maxima)代表了湿地生态生存策略的重要例子,即早春缺氧的季节性耐受。尽管在耐缺氧方面存在季节性变化,但G. maxima能够在竞争中胜过更耐缺氧的物种,如Filipendula ulmaria (Studer-Ehrensberger et al., 1993),这是因为它在早春生长的能力使其能够抢占其他后期发育物种的场地。在夏季,G. maxima根状茎不像其他一些湿地物种那样耐缺氧,不能在长时间的深度洪水中生存。当

关爱*社区Filipéndula社区甘油社区
图8.21后退的Filipéndula ulmaria斑块与前进的Glyceria maxima林分的界面。注意土壤pH值、电导率和有机质的逐渐变化。尽管沿样条的这一段的性质逐渐变化,但优势植物物种之间仍然有一个突然的边界。将其与图8.22(见正文)进行比较。(经Studer-Ehrensberger等人授权转载,1993年。)
关爱*社区Filipéndula社区甘油社区
图8.21后退的Filipéndula ulmaria斑块与前进的Glyceria maxima林分的界面。注意土壤pH值、电导率和有机质的逐渐变化。尽管沿样条的这一段的性质逐渐变化,但优势植物物种之间仍然有一个突然的边界。将其与图8.22(见正文)进行比较。(经Studer-Ehrensberger等人授权转载,1993年。)
如果在夏季高温(22°C)下缺氧,根状茎会损失总量的50%非结构4天碳水化合物储备(Barclay & Crawford, 1983)。在这种情况下,缺氧不耐受的根本原因是能量饥饿。
相比之下,越冬的根状茎在实验室中22°C下可以存活3周,在野外条件下可能存活更长时间。这种耐缺氧的季节性依赖的生理基础还不完全清楚,但部分原因可能是冬季碳水化合物储备高,而越冬器官的代谢活性较低。在芦苇枯死与异常高的矿物质养分浓度有关的地点,大甘油可以作为一个成功的替代物种。在这种看似有植物毒性的地方种植,从芦苇的衰落可以看出,大甘露的插枝可以在几天内产生新的根。在湿地环境中,大蕉的生存策略是一种逆境躲避策略,而不是一种逆境耐受力策略。的
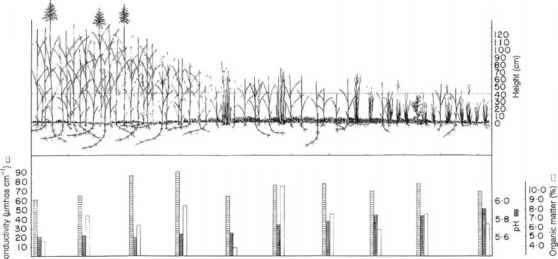
甘油群落黑苔草群落
图8.22大甘油和黑苔草的界面。注意沿样条的这一段土壤pH值、电导率和有机质的微小变化,优势植物物种之间没有突然的边界,并将其与图8.21进行比较(见正文)。(经Studer-Ehrensberger等人授权转载,1993年。)
甘油群落黑苔草群落
图8.22大甘油和黑苔草的界面。注意沿样条的这一段土壤pH值、电导率和有机质的微小变化,优势植物物种之间没有突然的边界,并将其与图8.21进行比较(见正文)。(经Studer-Ehrensberger等人授权转载,1993年。)
-
 invading a lake margin in Switzerland where it has replaced former vigorous stands of the common reed (Phragmites australis).")
- 图8.23入侵瑞士湖岸的甜菖蒲(Acorus calamus)取代了以前生机盎然的普通芦苇(芦苇芦苇)。
 invading a lake margin in Switzerland where it has replaced former vigorous stands of the common reed (Phragmites australis).")
物种有强大的能力将氧气从芽转移到根,发育良好的通气组织通过存在纵向排列的细胞的细分化来防止意外的水淹。这种布置可以防止在发生任何意外物理破坏时整个气陷层被淹没,因此降低了地下器官突然缺氧应力发展的风险(Armstrong et al., 1994)。然而,该物种的洪水耐受性的能力是严格限制的,在地下水位波动较大的地区,该物种不是芦苇的合适替代品。大甘油菊的短而软的叶子很容易腐烂,因此不能为长时间的淹没提供一个通气管。
8.7.4芦苇(芦苇芦苇)
芦苇(芦苇芦苇)与菖蒲和互花米草(同样是湿地土生植物)一样,一年四季都能忍受缺氧,并在生长季节有很强的能力将氧气从嫩枝输送到根部。因此,这些物种可以被认为是逃避者和杠杆式比长仪湿地生境缺氧胁迫条件的研究。然而,尽管芦苇和互花米草对淹没胁迫具有双重耐受性,但它们也会受到硫化物水平升高的影响(Bradley & Morris, 1990)。高硫化物浓度,在约1mm以上,经常出现在还原沉积物湖泊富营养化污染地区和河口淤泥。对芦苇耐硫化物的广泛研究表明,在低氧(<0.6 ppm氧)条件下,硫化物主要通过使金属酶失活而严重影响芦苇根系能量代谢,其枝条发育较弱。通常情况下,由于根茎表面加厚,硫化物不能直接进入根茎组织。然而,芦苇甲虫(Donacia claviceps)的幼虫啃食会造成损害,导致根状茎的空气空间被富含硫化物的水淹没(Ostendorp, 1993)。硫化物可以被转移到根茎中,在根茎中通过形成谷胱甘肽而部分解毒。中间化合物为半胱氨酸和谷氨酰半胱氨酸,o -乙酰丝氨酸作为硫化物受体。根状茎对硫化物中毒的敏感性低于根,但解毒能力有限,硫化物积累。在硫化物中毒的根中观察到以下现象:(1)腺苷酸能量电荷和总腺苷酸降低,(2)酒精脱氢酶活性降低,(3)缺氧后呼吸能力降低(Fiirtig等,1996)。
8.7.5两栖树木
树干粗壮、根系深厚的树木是在湿地栖息地承受缺氧的终极挑战。因此,令人惊讶的是,在世界上的许多地方,洪水对树木的生长没有损害。森林覆盖的低地密西西比盆地在美国,路易斯安那州和南卡罗来纳州的沼泽,以及热带沿海地区的红树林,都是树木生长能力的证明,即使在长时间的淹没是不可避免的。与所有其他高等植物物种一样,沼泽森林的树木有一个忍受持续淹没的时间上限,这是由组织更新所需的氧气决定的。
北美低地森林的树木也依赖于间歇性的低水位来再生。对于沼泽柏树来说,在许多树被杀死之前,可能需要连续四年或更长时间的淹没(Ewel & Odum, 1984)。在过去,每20-30年发生一次就足够了。然而,改进的水位管理已经消除了河流水位偶尔下降的数量,结果是在几个地区,如密西西比盆地和路易斯安那州的沼泽森林,这些森林的再生受到严重威胁。即使是沼泽柏树(Taxodium distichum)和水柏树(Nyssa sylvatica)等耐洪水树木的招募,在持续的洪水没有缓解时也会被阻止(Conner et al., 1981)。同样,据预测,莱茵河上更高的洪水水位将减少该河泛滥平原森林低洼地区硬木树种的建立(Siebel等人,1998年)。
继续阅读:湿地地点的热带与温带树木
这篇文章有用吗?


