方面的Highaltitude栖息地
高海拔环境地理因素,从气象学角度看,地质历史上。尽管这种异质性,他们仍然有某些共通的特性。在山顶一般最冷的栖息地在他们的特定区域,容易侵蚀(图10.9),较差的土壤和广泛的昼夜温度波动以及接触强风和高紫外线辐射,在温带,北方和北极区短生长期。适应问题的不断交替冰冻的夜晚之后提出了强烈的阳光,要求更大的生理耐受极端的温度会比在大多数开花植物是正常的。
在高空大气中水蒸气含量低,这显然干燥因素选择干旱防树叶高山物种。低温和石头覆盖的土壤高山然而保护土壤水分的蒸发,干燥的大气条件的补偿因素(见第三章)。
高海拔位置是否帮助植物避免危险的食草性仍然是一个悬而未决的问题。的植物进化限制气候和土壤条件下在山顶上与压力宽容分享相关的特征,即经济增长缓慢、扩展长寿、资源限制和低食草动物的适口性。污垢的C-S-R理论(污垢,2001)可以预见,删除重复的生物质食草动物会威胁到植物在高山环境中持久性(见章节3.4.2)。测试这一假说在高山的人群毛茛属植物(毛茛属植物glacialis)在一个高度的横断面奥地利阿尔卑斯山脉中部发现,15%至26%的r . glacialis植物在每个人口的迹象放牧损伤主要由雪田鼠(草原nivalis)(吴廷琰,1996)。只有很少的人口,孤立的冰川,在最高的网站(3310米)没有食草性的痕迹。定量评估的食草性的程度在两个高海拔(2600米和3180米)网站显示有相当大的损害:平均近25%的植物的总叶面积被一年以及65 - 85%的开花植物花序移除。尽管这些损失的大小生殖投资和每个植物叶子的数量开始改变明显在随后的一年。
一直声称,随着r . glacialis人口和其他类似擦伤了物种(如胚胎digyna)能够支持食草动物的种群alti-tudinal限制植物生长,没有明显减少的活力,那么这些植物不能被视为配件stress-tolerator,竞争对手和杂草方案提出的污垢(见章节3.4.2)。的污垢理论断言,不得不忍受极端的压力和干扰与生存不兼容(吴廷琰,1996)。然而,这种异议可以反驳的事实冰川灯芯草和类似高空物种并不是强调高海拔的网站。甚至有穴居的可能性,由小型microtines放牧可以提高植物多样性在高空网站通过增加网站的再生。
的高原牧场TransHimalaya,牧民认为小型哺乳动物作为他们的牲畜的竞争对手,造成草地退化,在许多地方积极消除植被的很大一部分。研究了小型食草动物如鼠兔的影响(例如Ochotona首要的,美国鼠兔)和田鼠(草原spp)发现,土壤扰动由于小型哺乳动物与高等植物多样性不会引起任何总体植被急剧下降(Bagchi et al ., 2006)。
许多高空物种是克隆,这可以提供稳定提供储备的高山草地(Erschbaner et al ., 2003)。似乎因此高空网站不是躲避食草性,尽管的低增长高海拔植物他们仍然拥有足够的

储备恢复甚至频繁放牧在这些严酷的暴露,甚至可以受益再生景观从穴居食草动物造成的干扰。
物理干扰山随地质。的大量浪费栖息地的增加而越松散岩是火成岩沉积岩,减少。扰动的侵蚀,然而,可以有好处和缺点植物生存。尽管侵蚀破坏植被还提供了新鲜的网站和营养物质,促进殖民和增长。一个鲜明的对比效果可以看到当比较山植被的硬山岩石加里东系统在挪威,苏格兰,纽芬兰的柔和的沉积岩新西兰的南阿尔卑斯山(图10.9)。在坚硬的花岗岩和变质苏格兰的岩石山坡上的土壤淋溶和营养不良,而较低的山坡上冲和富含营养物质从上面冲下来。相比之下,新西兰的气候hyperoceanic, fast-wearaybet雷竞技最新thering脆性岩石提供可用营养上地区的山脉。然而,在与苏格兰的石头的例子硬高降雨和快速冲蚀率在新西兰经历了迅速洗好的土壤和养分从山谷底部只留下gravel-filled流域(图10.10;参见图11.30)。
10.3.1地质和山组成
非常显著的地质对山地植被的影响已经注意到自从开始高山植物的系统研究。1749年的一篇论文提出了通过h . f .链接哥廷根大学的资格Goettingensis标本sistens veget-abilis盛宝calcareopropria,描述,可能第一次钙质和硅质岩石的植物之间的区别(沃尔特,1960)。最初,这个生态区别解释为是由于物理,而不是化学岩石之间的区别。石灰质岩石天气产生土壤温暖干燥,而基于硅质岩石土壤寒冷和潮湿;区别是一个重要的歧视性的特性在大多数山组成。更特别的植物有不同的反应,土壤化学博士在酸性土壤铝的影响(与)变得越来越可溶pH值低于5的食品。物种生活在酸性土壤称为嫌钙植物能够吸收或排除潜在的植物性毒素的铝离子,铁和锰的溶性减少亚铁和状态锰离子。
钙生植物植物生活在土壤pH值高(> pH值7.0)逃生的潜在毒性与价,Mn2 +这些只是溶于低pH值的土壤。作为螯合钙生植物缺乏特点嫌钙植物物种无法生存在酸性土壤。适应似乎是相互排斥的。不含铁和高效的手段
-

- 图10.10上水域Waimakariri河,南岛、新西兰、说明广泛沉积砾石侵蚀从脆弱的南阿尔卑斯山山脉。

表10.1。欧洲高山开花植物的例子作为对比存在替代物种(钙生植物和嫌钙植物)分别发生在基本和酸性土壤
钙生植物
蓍属atrata苔属植物curvula ssp。果Eritrichium nanum ssp。jankkae龙胆acaulis ssp。clusii Hutchinsia阿尔Minuartia弗娜报春花耳廓毛茛属植物alpestris杜鹃分子柳树婆罗虎耳草属aizoon虎耳草属oppositifolia景天属植物atratum Sesleria caerulea硅宾uniflora ssp。佩特拉- Soldanella阿尔
嫌钙植物
答:moschata curvula ssp。curvula
大肠nanum
g . acaulis ssp。kochiana
h . brevicaulis m . sedoides物种
毛茛属植物glacialis杜鹃ferrugineum柳树herbacea虎耳草属子叶虎耳草属rupestris景天属植物montanum Sesleria disticha硅宾rupestris
Soldanella pusilla
来源:各种各样的,包括沃尔特(1960);Reisigl &凯勒(1994);杰明(2005)。

图10.11杜鹃ferrugineum,嫌钙植物高山玫瑰。也称为生锈的高山玫瑰由于iron-laden毛发的势头远轴的叶片表面(见插图)。的去除铁在leafpresumably作为一种有效的方法,以避免过度积累有毒叶子中的二价铁。
图10.11杜鹃ferrugineum,嫌钙植物高山玫瑰。也称为生锈的高山玫瑰由于iron-laden毛发的势头远轴的叶片表面(见插图)。的去除铁在leafpresumably作为一种有效的方法,以避免过度积累有毒叶子中的二价铁。
锰离子嫌钙植物物种阻止他们生理上访问这些离子的必要的数量。这一起高碳酸氢的影响使他们的叶子变成萎黄病的——条件通常称为lime-induced萎黄病(马斯纳,1995)。
许多种和亚种不同基因密切相关,他们的偏好钙质和硅质土壤。这是特别明显的山阿尔卑斯山(表10.1)等基础地质和土壤之间的关系不戴面具的深层土壤覆盖泥炭或发展冰川沉积物。
最著名的例子之一是不同形式ofalpine杜鹃花,“高山玫瑰”,杜鹃ferrugineum发生在酸性土壤,而毛高山玫瑰(r .分子)是局限于石灰性土壤。甚至特定的网站如雪补丁不同物种的存在取决于底层的钙质和硅质岩石(无花果。10.11 - -10.12)。
相比之下,在最高峰,喜马拉雅山比高度、地质似乎不那么重要,降水和方面。可以通过从一个岩层到另一个没有任何明显的植被的变化。因此,long-leaved松(松果体roxburghii)生长在酸或基本岩石没有任何明显歧视(Polunin &斯塔特,1984)。
之间的差异的基础组成的酸性和钙质土壤可以部分原因是非常真实的物理差异钙质和硅质岩石的物理性质,以及土壤的化学性质。
-
")
- 图10.12杜鹃花,钙生植物高山玫瑰。(图a . Gerlach博士。)
")
钙质土壤通常是温暖和干燥和硅质土壤是凉爽和潮湿。这与不同的反应土壤化学和植物生理学的pH值、钙和碳酸氢根离子导致植物分布明显的区别。这是特别明显的山组成,早期和晚期站点之间高度的温度大幅限制区分为重启增长在春天。因此,有一个显著的物候维度温暖干燥石灰性土壤的优点和缺点而不是通常凉爽和湿润的环境与硅质土壤。
10.3.2绝热递减率
的冷却效果增加高度绝热递减率的影响,这是消极的垂直温度梯度由通过周围空气的垂直运动吗流体静力学平衡(Calow, 1998)。当空气是被称为不饱和干绝热递减率和的值为9.8°C公里,提供移动和环境空气几乎相同的温度(通常是这样)。然而,如果保持饱和水蒸气的凝结在空气上升,然后释放潜热减少了绝热递减率(图10.13)。
在温暖的低对流层(地球大气层的最低部分和一个最天气现象发生),饱和adia-batic递减率可能只有一半的干绝热递减率盛行在寒冷的高对流层(Calow, 1998)。一个全球性的意思温度递减率5.6°C km-1已经报道了山(Korner, 2003)。然而,有很强的地方温度反演,沿着主要在格陵兰冰原的边缘,价值高达12°C km-1已经被记录在案。混合的不同的空气质量可以修改的吗温度变化与高度。海洋区域挪威和苏格兰西部的天气由极地海洋空气经常体验。这气团,冷冻了格陵兰冰帽在较低的水平,将温暖传递到北大西洋漂流。然而,上面的空气质量仍然冷。低水平的混合热空气和高水平冷空气带给这些土地接壤的东部海岸的北大西洋更快速下降温度与高度通常比预测。在苏格兰西部,降低温度8 - 9°C km-1高度并不少见。
生态,这些不同的绝热率产生深远的山地植被区划的影响。hyperoceanic条件的苏格兰北部和西部群岛山区植被成带压缩由于绝热率高。有可能在奥克尼群岛(59°N)站在山区苔原植被类型在峰会上只有420 m,并查看低于海平面大麦作物生长(图10.14)。
10.3.3山地地形和生物多样性
任何特定的潜在植物区系多样性山通常可以预测在距离其形状,地质学和岩石结构。尽管可能出现令人生畏的前景植物殖民经常奇怪多少高山支持丰富
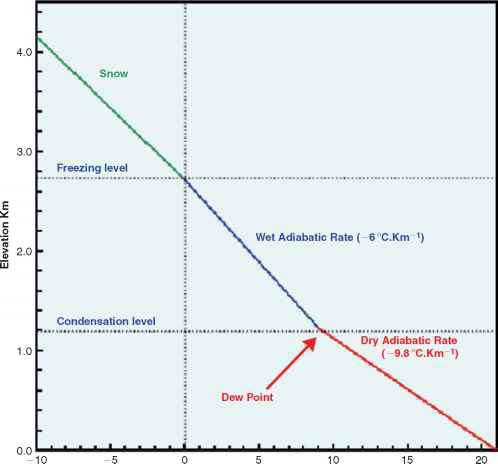
图10.13绝热递减率,高度的函数。(基于特拉,1963年,页246 - 252)。
温度°C
图10.13绝热递减率,高度的函数。(基于特拉,1963年,页246 - 252)。
高山植物。斜坡石堆和梯田看起来不稳定,但他们可以提供,并根据方面地质构造,多变和有利的栖息地,是足够的一系列适当改编的物种。将军山地形粗糙度也使丰富的微型网站,局部环境条件可以提供温度制度,可以抵消海拔和纬度的影响。
山脉和他们的方向也很重要的生物地理学为移民提供范围,特别是气候变化重要时期。raybet雷竞技最新南北方向通常被认为是最有利于改变温度。然而在东西方向
从海洋到欧洲阿尔卑斯山可以适应变化大陆气候raybet雷竞技最新。毫不奇怪人们往往可以预测在许多高山植被的多样性取决于山可以被描述为凹或凸。凹山提供水库土壤、水和营养物质,而凸山区往往缺乏这些资源。因此描述符的曲率和粗糙度解释物种分布的变化比“古典”地形属性,如海拔或接触(Gottfried et al ., 1998)。
消极的特性孤立的山在一定程度上抵消当山既大又高。相反,在小山脉,
-

 illustrating the rapid change in climate and vegetation with increasing altitude in oceanic climates. Note the tundra vegetation in the foreground and active agriculture in practice only 305 m (1000 feet) below.")
- 图10.14从病房希尔,霍伊,奥克尼(59°N)说明气候和植被的快速变化和增加高度在海洋气候。raybet雷竞技最新注意苔原植被在前台和活跃的农业实践只有305米(1000英尺)以下。

 illustrating the rapid change in climate and vegetation with increasing altitude in oceanic climates. Note the tundra vegetation in the foreground and active agriculture in practice only 305 m (1000 feet) below.")
生物气候植被成带压缩(低、中、高的高山,参见10.5.1)与更大的山脉。
德语生态学家将这一现象称为Massenerhebung或Massenerhebung effekt(质量海拔效应),山的大小可以改善自然环境。效果是最明显的大型山脉聚集在一起。中心等温线水平升高,并创建一个更大陆性气候,通常与云量减少,导致raybet雷竞技最新积雪区雪(永久)
水平撤退到一个更高的高度。的高山植被因此可以发现在大型山脉高海拔比小山脉。前面的范围,然而,不存在很大差异较小的山脉。
由于这种Massenerhebung效应也有更大的生产种子在高海拔地区,它提供了一个更大数量的繁殖体吹洗沿着这反过来增强了高山植物在海拔较低地区的存在(Ellenberg, 1963)。
继续阅读:分带的案例研究
这篇文章有用吗?
读者的问题
-
abbondio3个月前
- 回复
-
考3个月前
- 回复
-
贾米拉•3个月前
- 回复
-
codey3个月前
- 回复
-
蒂娜3个月前
- 回复