生活史策略
当更具侵略性的物种因环境变化或不确定性而无法达到最大生产力时,竞争力较弱的物种的生存能力长期以来一直被认为是物种多样性的基础。人们已经做了许多尝试,以寻找各种适应或策略的组合,以描述不同物种对资源竞争压力或邻居干扰的反应方式。“战略”一词,当应用于生态学时,与它更常见的军事用法分离开来,仅仅表示“涉及大量相互作用和共同进化特征的适应的综合体”(Calow, 1998)。最近,植物生命策略(定义见表3.2)在生态学思想中获得了新的重要性,因为它们可以形成概念化维持生物多样性过程的基础(见第2章)。然而,当这些分类被应用时,例如在植被对变化环境响应的计算机模型中,重要的是要记住,就像竞争的概念一样,它们只是人类用来从植物对环境的无限多样的行为反应中创造秩序的感知,因此它们是想象的场景,而不是生物现实。
3.4.1两类人生策略
最早的生活策略分类是基于对资源使用的两类划分,使用诸如资本家和无产者等术语,这可能反映了这一时期激进的政治思想(Grime et al., 1988)。这种和其他双向分类在本质上预测了r和K选择策略的分类,其中生物的划分是基于它们的生活方式是否适应频繁干扰的环境或生态位,或者它们是否能够持续开发高产的栖息地(图3.23)。在选择的物种中,典型的栖息地是资源是短暂的和暂时的,因此增长最快的种群占主导地位。这样的群体通过增加出生率(幼苗产量)或更有效地减少世代时间来增加其内在人口增长率。这种快速增长的种群死亡率很高,特别是在幼苗阶段。因此,有效的传播机制将是有利的,因为它们将通过将种子分发到可能有利的地点来降低幼虫的死亡率。杂草物种有时被描述为典型的r-选择物种。然而,这并不是一个普遍的概括。许多杂草品种具有很强的竞争性,并表现出一定程度的生态类型灵活性,使它们能够适应不同的情况,并运行自己的连续序列。对于危险的多年生杂草,如匍匐蓟(Cirsium arvense)、野旋草(Convolvulus arvensis)、多年生南蓟(Sonchus arvensis)、麦草(Elytrigia repens)、蕨菜(Pteridium aquilinum)和野马尾(Equisetum arvense),情况尤其如此。
长期稳定的栖息地承载的物种种群通常接近承载能力(K),种群进一步增长的空间很小。这样的
表3.2。植物按功能分类的定义示例
物种的生命形态分类:根据与其环境相关联的形态特征对物种进行分类。Raunkiaer(1934)的方案,基于多年生芽在不利季节高于地面的高度,仍然是应用最广泛的系统。
生命史特征生物体生命周期中与出生率和死亡率最直接相关的特征,从而与达尔文适应性最直接相关。生活史特征可以被视为个体分配其有限资源的方式(Calow, 1998)。
生活史策略在物种或种群中广泛出现的相似或类似的生活史特征的分组,使它们在生态学中表现出相似性(Grime, 2001)。
功能类型对一套环境条件有相似反应的物种或物种群。在实践中,这些分组通常与生理功能有关,因为它受到不同环境的影响。
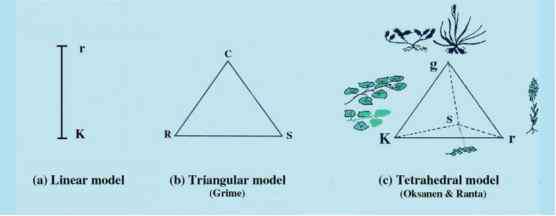
图3.23解释植物对竞争和环境胁迫反应的2、3和4种分类策略。(a) r和K之间的基本两种分类体系选择。(b) Grime在竞争对手(C)、应力耐受力(S)和粗骨料(R)之间的三角分类模型。(C) oksanen和Ranta, 1992的四面体修饰,表面有三个主要策略(K、R和g)四面体.g策略是典型的ericoid和禾草状的具有低根枝比和低身材的种。额外的策略位于前平面后面,代表支持组织的分配减少,极端类型是肝脏。(经Oksanen & Virtanen许可转载,1997年。)
图3.23解释植物对竞争和环境胁迫反应的2、3和4种分类策略。(a) r和K之间的基本两种分类体系选择。(b) Grime在竞争对手(C)、应力耐受力(S)和粗糙体(R)之间的三角分类模型。(C) oksanen和Ranta, 1992的四面体修饰,在四面体表面采用三种主要策略(K、R和g)。g策略的典型是低根冠比和矮个子的ericoid和graminoid种。额外的策略位于前平面后面,代表支持组织的分配减少,极端类型是肝脏。(经Oksanen & Virtanen许可转载,1997年。)
物种可以有较低的繁殖力水平,这被较长的寿命所抵消。对特定竞争策略的选择使得K-selected物种高度专门化,从而对环境变化更加敏感。它们也不太能从低人口密度中恢复过来,因此很可能是濒临灭绝的物种。无性繁殖几乎可以使人永生,从而提高生存的可能性。这种情况在北方的许多白杨种群中都可以看到,这些白杨在小冰期(大约持续在13世纪到19世纪之间的一段冷却时期)很难结子,但由于吸吮的能力而幸存下来。同样,在森林再生的主要边界之外的树木也通过分层来维持。一些外来物种由于其营养繁殖的能力而达到害虫的地位。在苏格兰,常见的杜鹃花(rhododendron ponticum)被引入到高地,由于它的分层和能力树荫下公差能胜过许多原生林地植物群。
3.4.2三级生活策略
尽管简单的二元划分足以突出植物和动物竞争行为中一些最鲜明的二分法,但当通过三策略模型来观察植物竞争时,情况有了相当大的改善。
人们还提出了多种植物行为的第三类。逆境选择(Southwood, 1988)使栖息地的质量发挥作用,在这种环境中,生物体可以以代谢代价适应应激条件,但会导致生长或繁殖受损。Grime提出的竞争策略的第三维度概念认识到幼株与成虫和成熟植物表现出的行为之间的死亡率差异。再生策略的变化利用了资源投入、流动性和休眠的差异,这些差异主要表现在再生阶段。因此有人提出(Grime et al., 1988),有两类主要的外部因素影响植物的生命策略。第一种可能被描述为压力,包括所有可以限制光合作用产生的现象,如缺乏光、水和矿物质营养素或温度过低。
第二类被称为干扰,包括导致生物量部分或全部破坏的所有现象,如草食、践踏、割草引起的人类干扰和风损害、霜冻引起的自然物理干扰干旱、水土流失和火灾。在三类生命策略概念中,植物只能处于两种极端边缘条件,即极端胁迫或极端扰动。Grime认为,极端压力和极端干扰的结合导致了不适合植物居住的地区。
3.4.3四类人生策略
然而,在某些情况下边际的栖息地这促使人们认为,将植物想象成有多达四种基本的生命策略可能更为现实。在北极、亚北极和山区的一些栖息地,有可能在资源严重有限的环境中进行高强度的放牧并生存下来(见第10.3节)。在许多情况下,甚至可以证明,这种放牧对开花植物的生存是必不可少的,以避免凋落物和阴影的积聚,并在土壤中形成致命的冰楔。
即使在相对贫瘠的北极和高山栖息地,放牧也一直存在,并经常伴随着寒冷扰动和侵蚀的强烈物理干扰。有人认为,放牧选择的特征与破坏性的物理干扰完全不同,后者没有幸存者(Oksanen, 1990,1993)。地上组织的损失显然是为了选择高根枝比和通过水平根状茎进行营养繁殖,特别是如果损失是由食草动物引起的,它们更喜欢营养丰富的花芽。此外,草食性选择坚韧、狭窄、细裂或鳞片状的叶子,而反对宽阔、中胚性的叶子。放牧对后者尤其有害,因为部分食用宽的中胚性叶片会造成长伤口,使剩余的组织暴露于干燥和寄生真菌的入侵。
植物已经进化出某种稳定的防御策略这一概念表明,在抵御放牧方面有两种主要策略。植物要么将大量减少的碳投入防御性化合物中,这不可避免地导致低生长速度,要么仅通过现成的手段来保护自己无机物质(如二氧化硅)或通过机械威慑(如厚角质层,刺),即使在低资源投资水平也相对有效。在高强度放牧中生存的两种主要方式被描述为ericoid和graminoid策略(Oksanen, 1990)。奥克萨宁认为,典型禾本科植物的主要优势——它们能够从基部的花萼分生组织迅速产生直立的叶子——在持续的、激烈的放牧的栖息地中会减少,在那里,芽的竞争从未变得激烈。以上四类策略的性质可以通过将Grime的三角形策略图扩展为四面体来相互比较(Oksanen & Ranta, 1992)。该方案的一个方面代表了k策略或竞争力感知Tilman (Tilman, 1988),即一套形态、生理和繁殖特征,使植物能够在关键资源被这些植物压低到低水平的环境中完成它们的生命周期。另一个极是经典的R策略,它可以被视为与Grime的R或粗糙策略相同。Grime的竞争者或c -战略家被认为是介于K-战略家和r-战略家之间,如Grace (Grace, 1990)所提出的,它们适应于资源丰富、干扰竞争盛行的环境。第三极由g-战略家组成,适应地面组织频繁但微小的损失。s策略表示减少支持组织的分配,极端类型是肝脏。该系统还设想了ericoid、graminoid和Dryas策略的进一步细分,它们是g-策略更一般概念的子集(Oksanen & Virtanen, 1997)。
尽管其概念复杂,与传统的两类和三类策略概念相比,这种对植物生命策略的四类分类的讨论有助于证明,随着条件变得越来越边缘植物生存在高等植物中,这并不必然伴随着任何形式或功能多样性的丧失。正如这本书中所论述的那样,在边缘地区生存的植物并不一定是因为它们适应环境的多样性而贫瘠。
继续阅读:混合区
这篇文章有用吗?
读者的问题
-
Gianna25天前
- 回复
-
哈利1个月前
- 回复
-
KEIJO2个月前
- 回复
-
Mehret2个月前
- 回复
-
Efrem3个月前
- 回复
-
裘德3个月前
- 回复
-
桑德罗FANUCCI3个月前
- 回复
-
莎拉3个月前
- 回复
-
dawit bisrat3个月前
- 回复
-
玛丽亚(女名)3个月前
- 回复
-
比尔3个月前
- 回复
-
曼迪3个月前
- 回复