脆弱的海洋沉积生态系统服务对人类活动
保罗稳压器Snelgrove Melanie c·奥斯汀史蒂芬。霍金斯,托马斯·m·Iliffe罗纳德·t·Kneib丽莎·a·莱文Jan戈Weslawski罗伯特b . Whitlatch,和詹姆斯·r·Garey
海洋沉积地球表面的生态系统包括超过任何其他栖息地,但许多人认为海底是一个巨大的、单调的环境远离人类的干扰。生物多样性是通常认为的后果很小的资源我们从海洋中提取,海洋环境的健康和人类的生活质量。尽管如此,渔业等海洋沉积物提供重要的可推断出的货物。他们也在全球扮演监管角色转移和原材料和能源循环(见第4章Weslawski et al .)。许多生态系统的过程(美国查宾et al . 2002年)发生在海洋沉积物中也有重要的影响人类社会的可持续发展的生态系统服务价值(如海岸线稳定,废物回收等)。海洋沉积物的高度可见的海岸线(图7.1)远程和鲜为人知的深海不同风险威胁和伤害威胁如果暴露的概率;这两个问题导致的脆弱性,在这里定义为生态系统遭受伤害的倾向暴露在外部压力和冲击(墙et al . 2001;Folke et al . 2002年)。本章的目的是检查潜在威胁生物多样性的海洋沉积的生态系统。我们问生物多样性的变化是否会增加系统的脆弱性的流程和服务,我们检查他们的潜在恢复的影响。在大多数例子中,损失的服务提供的海洋沉积物尚未解决的科学家;讨论,因此,关注过程与底层暗示这些影响服务(见Weslawski et al .,章节
-

- 图7.1。示意图总结的主要威胁生态系统货物和服务在河口和沿海生态系统。

表7.1。总结主要威胁海洋沉积系统和它们的尺度体现。
规模
问题
生态系统影响最大
小范围内
(个人湾)
中期(区域)
大规模(盆地规模)
入侵物种疾病沿海开发/栖息地改变hydrologie改变过度捕捞和栖息地的破坏eutrophica/污染气候变化,包括:海平面变raybet雷竞技最新化的降雨模式风温度和循环河口盐度紫外线辐射,河口湿地,湿地湿地、河口湿地、河口,潮间带书架,斜坡,河口,河口湿地,货架湿地、潮间带、河口湿地、潮间带书架,河口潮间带,湿地、潮间带
个人河口货架河口系统区域
斜率深海地区地区
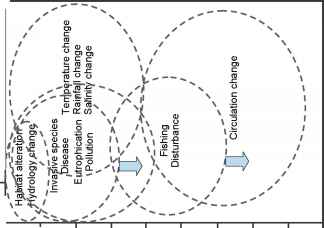
图7.2。尺度的威胁到海洋沉积的栖息地。圆圈表示疫区和栖息地的范围。箭头为入侵物种/疾病和渔业干扰表明潜在的大尺度与人为干扰增加的威胁。
4)。在第四章的体积,海洋系统分为河口,大陆架和深海沉积物。河口包括沉积栖息地在陆海交界海水由淡水输入测量稀释,conti-nentalshelf指之间的海底大陆和大陆坡的顶端(~ 130米深),和深海栖息地包括相对陡峭(~ 4°)大陆坡,从大陆架的边缘延伸到大陆(4000 ~),成绩上升到巨大的深海平原(4000 - 6000),覆盖大部分的深海底。海山是水下山脉深海平原上扩展几千米的海底。
威胁和尺度的脆弱性
沉积生态系统面临多种威胁源于人类活动(表7.1),可能对物种组成产生重大影响和生态系统过程在当地(湾或半封闭的沿海地区)、区域(数百公里)和流域尺度上阶段(钟et al . 2001年)。像其他生态系统、海洋环境承受能力,从人为干扰中恢复过来。例如,可持续渔业规划设计是有可能的,因为popula生物有一种天生的能力,增加他们的数量,和个人被钓鱼取而代之的是那些逃脱的渔业的后代。同样,许多海洋系统可以容纳一些污水输入没有生物多样性的丧失或代的缺氧。问题是,干扰往往超过系统恢复的能力,从而导致物种减少,在某些情况下,损失的生产商品和服务的能力。大小的系统及其邻近人类可能在不同尺度影响的脆弱性,因此我们检查漏洞的威胁的规模影响(图7.2)陆海交界的深海。
局部范围的威胁(数万公里公里)
河口更有可能比大陆架(轻轻倾斜的地区从0 - 130米深度之间的海岸线和大陆坡的上边缘)和深海(大陆架以外的地区,包括大陆坡130 - 3000米),大陆崛起(~ 3000 - 4000米),和深海平原(~ 4000 - 6000 m])系统经历严重的局部范围的影响,因为他们在空间较小的程度上,减少对相邻系统开放,和身体上更接近人类(Levin et al . 2001年)。然而,我们也认识到当地严重影响(例如,许多个人拖网)可以传播,成为区域(例如,大规模捕捞的影响)。下面我们轮廓局部范围的威胁。
外星人类群
外来物种通常介绍了从点光源(如船舶压载水和船体污染,海水养殖海洋水产养殖)活动;看到卡尔顿&盖勒1993),尽管在物种高度侵袭性的情况下,他们可以快速创建问题区域范围内通过快速传播的繁殖体(Wehrmann et al . 2000年)。入侵物种也可能表现出人口落后超过很多代成为问题之前(骗子&苏尔1999)。在河口栖息地,生物入侵排名仅次于生境改变之间的潜在威胁生物多样性(Vitousek et al . 1997;卡尔顿2001)和入侵利率沿海海洋系统加速了在过去的200年里(Ruiz et al . 2000;卡尔顿2001)。最底栖动物物种入侵的甲壳类动物和软体动物(Ruiz et al . 2000),但多毛类(Roehner et al . 1996),植物,病菌也改变了底栖生物多样性(卡尔顿2001)。介绍了植物可以改变社区和生态系统过程,因为他们在河口食物网营养的重要性,影响物理生境结构(如建筑、沉淀)。例如,芦苇南极光减少植物物种多样性,入侵(由Lenssen et al . 2000),降低土壤含盐量和水位,减少microtopographic特性,和改变沉积物氧化(温德姆&莱斯罗普1999),但它不是已知减少淡水湿地底栖动物物种多样性(Ailstock et al . 2001年)。入侵可以降低密度,提高物种丰富度,改变infaunal成分在潮汐湿地(Talley &莱文2001)和未知功能的后果。 Hybridization with local species can also alter or reduce genetic diversity (Ayres et al. 1999).
疾病
所有生物体疾病代表自然威胁。人类会加剧这种威胁通过生理应激对疾病的易感性增加,加速疾病的传播,或通过引入污染物进入海洋环境,以控制疾病。效果可能包括商品和服务的损失或变更的过程。例如,化学污染可能导致慢性病变(摩尔et al . 1997年)和增加寄生的影响(汗1987年)底栖鱼类,减少商业价值和潜在影响渔业的可持续发展。运输的有毒腰鞭毛虫囊肿在压载水会污染新领域(Hallegraeff & Bolch 1992),导致海水养殖和野生贝类产量损失。海水养殖中抗生素的使用似乎是一个本地化的威胁,但药物用于控制寄生虫可能对底栖无脊椎动物的毒性,远远超越mar-iculture地区(Goldburg et al . 2001年)。许多海水养殖项目也使用非物种或股票,并介绍或逃的病原体导致的外来物种入侵问题河口(卡尔顿2001)。然而,几乎没有证据表明这一新兴行业大规模疾病对底栖生物群落的影响(罗斯柴尔德et al . 1994;Naylor et al . 2000年)。
沿海开发和栖息地的改变
人类活动改变的沿海栖息地的物理结构在空间和时间尺度的范围。Lerberg et al。(2000)相关的数量和类型海岸线发展物种丰富度和种群的主要官能团的变化。也许最直接的负面影响infaunal(生物生活在沉积物)社区与沉积物扰动在航道疏浚导航(Newell et al . 1998年),和更多的慢性影响重型装备(如鱼拖网,扇贝疏通)经常用来收割河口和沿海的物种(摩尔HallSpencer & 2000;画眉&代顿2002)。甚至相对局部泥沙扰动(如管道安装,增加土地侵蚀径流的沉积物)会影响macrofaunal生物多样性(Lewis et al . 2002年)。这些干扰常常破坏或消除大,无柄,和长寿的底栖生物滤食无脊椎动物(如珊瑚、海绵、双壳类)、海草、红树林、沼泽植被。这样就降低了结构的复杂性和栖息地的多样性以及系统的能力陷阱沉积物(莫里斯et al . 2002),改善水质(科恩et al . 1999年),和缓解压力的影响,如缺氧(耗尽氧气浓度)(1998年Lenihan &彼得的儿子)。长须鲸笔海水养殖,缺氧下笔减少底栖生物多样性(韦斯顿1990)。干扰在自然栖息地包括河口经常创造条件,支持不同的物种比那些发生在安静的地区,与潜在影响人类生态系统过程和相关的利益。
alteration-construction频道的蓄水池、码头、道路和海岸线铠装(从侵蚀保护近岸产权)-影响的沿海栖息地的功能(Kneib 2000)。海岸线铠装,在防护物理结构,如混凝土防波堤保护沿海财产,防止自然湿地内迁的海平面上升(Pethick 2001),构成严重的长期威胁的多样性和生态系统过程(莫里斯et al . 2002年)。装甲海岸线硬衬底生物群(即支持。暴露的基础上发现的,生物和其他nonsedimentary栖息地)在以前支持的环境中沉积的生态系统(戴维斯et al . 2002年)。关键营养链接和生态系统过程(例如,能量流动)也取决于维持河口和栖息地之间的物理通道材料和生物体的运动(Micheli &彼得森1999;Kneib 2000)。
河口函数作为移动物种托儿所收获其他地方(例如,架子近岸水域)和有时幸免的直接影响商业(虽然不是娱乐)渔业。然而有许多过度开采资源河口和海湾的例子包括减少或灭绝的底栖生物filterfeeders,关键的官能团,强烈影响bentho-pelagic耦合(底栖生物和水体底部之间的联系(远洋)环境)和水质(杰克逊et al . 2001;代顿et al . 2002年)。
对地区级的核的威胁
(数百到数千公里)
水文变化
水文变化发生在河流改道或流出明显减少用于其他目的(如水力发电项目,灌溉)。效果通常是本地的(如单一河口)但重要的娱乐会影响整个沿海地区。增加水的开采和消费降低地下水位,减少淡水输入,增加海洋影响河口底栖生物多样性和生态系统带来严重后果的过程。淡水的数量或时间变化流干扰之间的平衡的影响陆地和海洋的河口环境(Sklar &布劳德1998),并影响物种的生命周期依赖于季节性周期淡水河口输入。地区降水与潮间带湿地和红树林的面积范围内温带和热带河口和沿海湾(Deegan et al . 1986年),和物种多样性在河口盐度的变化有关(Kalke & Montagna 1991;Jassby et al . 1995年)。脉冲洪水事件可以改变泥沙和养分输入支持deposit-feeders泥沙量的(高)和suspension-feeders泥沙量的(低)在不同季节(Salen-Picard & Arlhac 2002)。同样,减少海洋交换可以创建过度或hyposalinity和缺氧(泰斯科&里奇2001)。
海水养殖
研究大规模养殖对底栖生物群落的生物多样性的影响很少,但是环境的影响海水养殖操作与上面列出的对生物多样性的威胁,尤其是对栖息地的破坏。在许多发展中国家,潮间带湿地红树林和其雷竞技手机版app他已经被虾海水养殖池塘。在泰国,河口红树林的54%在1961年被转换为其他用途,到1993年,主要是为建设虾池(Macintosh et al . 2002年)。的高底栖生物群落的生物多样性和高生产野生虾和其他渔业的物种在自然红树林系统可能永久失去(Naylor et al . 2000年)因为试图恢复这些受损的栖息地很少生产中发现的甲壳类动物和软体动物的物种多样性不成熟的红树林栖息地(Macintosh et al . 2002年)。
过度捕捞和栖息地的改变
浅海岸和货架系统往往大量捕捞目标物种通过捕获与可观的间接伤害。目标物种的过度捕捞死亡率和/或捕获可以大幅改变食物网(保利et al . 1998年)。底拖网渔业也可以被均质化大面积破坏海底为底栖生物提供栖息地和表明物种(代顿et al . 1995年)。尽管过度捕捞是集中在河口和货架地区(南风et al . 1996年),它已蔓延至海底山脉和大陆斜坡。
渔业影响最大的担心在货架系统的威胁(国家研究委员会1995年)。在河口海岸和货架系统中,过度捕捞减少库存水平,改变营养支持流程和食物网动力学之间的远洋和底栖生物区也在海底2003年(保利& MacLean)。商业捕杀鱼类通常是顶级捕食者,和它们的过度开采级联效应通过降低食物链的部分(迈尔斯和蠕虫2003;蠕虫&迈尔斯2004)。同样,收获大滤食动物的双壳类消除人口底栖生物系统中执行关键过程(保利et al . 1998;杰克逊et al . 2001年)。食物链的破坏通过过度捕捞、富营养化和入侵物种一致行动进一步减少可用性的鱼类和贝类食物(兰斯洛特et al . 2002年)。
尽管休闲渔业可造成大量死亡的顶级捕食者(代顿et al . 2002),底栖生物多样性受到的影响研究较少,结果喜忧参半。有证据表明从操纵(Kneib mesocosm实验
1991;达菲&干草2000)表明营养相互作用涉及顶级捕食者可以级联影响底栖生物组合的成分(因此生物多样性),但这是不确定mesocosm结果可以扩大到任何大,开放系统。几乎是不可能的,调查这个问题在许多地区由于公众和政治阻力建立海洋保护区,禁止渔业开发的完全保护。例如,在美国只有不到1%的海洋环境不仅仅是名义上的保护;大多数都是珊瑚栖息地(所有2002)。
在更大的尺度上近海和沿海,栖息地的改变是由拖渔具通过沉积物,从而打乱了化学和生物梯度。全球拖网影响数千平方公里的海底大陆架,虽然效果取决于沉积物类型、渔具、拖网频率(牧羊犬et al . 2000年)。较大的大型生物,生活在和沉积物epifauna最容易受到拖网捕捞死亡率(Kaiser et al . 2000),受伤从渔具(拉姆齐et al . 1998年),和接触捕食者。海螺等软体动物身体滚被钓鱼齿轮和暴露,因此,更容易受到捕食(拉姆齐&凯泽1998)。食腐动物食物摄入增加捕捞区域(Kaiser &斯宾塞1994),和鲨鱼和鳐鱼把拖网渔船和食品(史蒂文斯et al . 2000年)。更大的海底动物和epifauna创建本地,小规模的栖息地破碎通过改变水流或创建生物结构如洞穴和管,或通过移动沉积物和喂养。通过减少这些数量,拖网同质化沉积物和景观互连通过生境斑块和聚集地(南风et al . 1995;画眉et al . 2002年)。生物体创建小规模的异质性和栖息地结构也稳定沉积物通过吸积和水流在湿地中接口的变更。
大深度的深海平原,地处偏远,收获的物种丰度低的集中大部分深海渔业活动上大陆坡中期的深度。深海生物通常生长缓慢,在稍后的年龄比浅水物种、成熟和生产相对较少的后代(梅里特& Haedrich 1997)。捕获率下降和股票大小已经观察到的商业利用鱼类克拉克(1995年),深海蟹,虾(Orensanz et al . 1998年)。附带捕捞也引起丰富的深海鱼类数量的急剧减少,没有针对性的渔夫在北大西洋(Baker & Haedrich 2003)。在其他系统中,捕鱼删除虚拟的顶级捕食者发挥监管裁剪作用在深海多样性维护(代顿&赫斯勒1972;迈尔斯&虫2003)和补丁创造(Grassle Sanders & 1973)。渔具可以有显著的影响类似于深海海底栖息地在货架上看到环境(Koslow et al . 2000年)。和深海珊瑚海山(Lophelia)珊瑚礁是这个问题的一个极端的例子,在破坏性的拖网齿轮损害epifauna特有的特定的海底山或礁区域和可能提供关键的鱼类和其他生物的栖息地(Koslow et al . 2001;Hall-Spencer et al . 2002年)。
更均匀的深海设置,栖息地的破坏和掠夺者的影响去除由绝大程度上减轻环境。渔业对深海多样性的影响(与高水平的特有现象除了海山)是很难知道的,考虑到物种总数的估计存在不同的一个数量级(Grassle & Maciolek 1992;Lambshead &鲍彻2003)和分布地图不存在对大多数深海物种。
删除远洋高级消费者(如鲸鱼,金枪鱼)和钓鱼的食物网也可能级联影响深水食物链(Butman et al . 1995年)。输入大量有机质下降到深海楼可能减少,影响粮食供应许多食腐动物物种,导致其他深海鱼类的饮食。
深海拥有丰富的矿产和石油资源。锰结核(如磐石般坚韧,高尔夫球大小的结构,含有丰富的锰和其他矿物质和密集的浓度在某些领域中发现的深海平原)和外壳,poly-metalic硫化物(chimney-like存款形式在深海热液喷口),磷灰石都含有有价值的钴、镍、铜和锰,但他们的提取还不是经济可行的史密斯(Glover & 2003)。潜在影响底栖生物在一定程度上取决于矿业废物(主要是沉积物)在海底排放。任何场景包括从采矿设备损坏和破碎,但海底放电也可能抑制生物。减少一些效果包括初始增长紧随其后megafaunal丰度主要由清除物种(布鲁姆et al . 1995年)和没有改变或减少macrofaunal丰度和多样性(博罗夫斯基2001)。尽管底栖生物种群可以从模拟扰动在三年之内,恢复多样性影响可能保持七年后。实验表明相对本地化,noncatastrophic影响,虽然这些实验表明谨慎的小规模商业影响推断(Ozturgut et al . 1981年)。尽管如此,专门的锰结核动物群,可以减少或消失在激烈的采矿方案(泰尔et al . 1993年)。
石油和天然气钻探在架子上打破已经从只有一种可能性,半个世纪前,在近几十年来成为现实。例如,巴西公司PetroBos一些生产活动深度大于1800 m西非(史密斯Glover & 2003),和探索其他公司发生深度大于3000 m在墨西哥湾(矿产管理局2004)。如果仔细进行钻探,多样性和生物量的影响可能非常本地化(公里范围),除了大的影响与重大泄漏(泰尔2003)或长时间尺度上钻探。相比之下,甲烷水合物的开采,虽然还没有在技术上可能,可能破坏边坡区域,导致质量下滑希尔与大规模死亡事件(2003)。甲烷水合物是一种气体,冻结深度大于300 m,并可能有一天是最重要的深海海洋资源,因为它认为掌控两倍碳甲烷水合物和其他来源ios版雷竞技官网入口 (美国地质调查局调查简报:http://marine.usgs.gov/fact-sheets/gas-hydrates/title.html)。
富营养化和污染物
河口和半封闭的海湾经常支持密集的人口和工业,相关高污染物的输入(如重金属、碳氢化合物)和营养物质,这可能会导致浮游植物大量繁殖,随后衰变和创建底部缺氧。河口和海湾有限的交流与开放海洋非常脆弱;然而,大规模的缺氧事件越来越频繁地对大陆架区域附近的输入来自新河口或大型河流系统(迪亚兹和罗森博格1995)。尽管许多河口和架子上的生态系统有着显著的回收有机物和营养物质的能力没有引起缺氧,平衡社区的微生物和bioturbators需要提供这个生态系统服务。
污染物输入河口和海岸从和面源是越来越全球问题点(伯施et al . 2001年)。许多河口接触到广泛的污染物,包括石油泄漏导致底栖无脊椎动物复杂变化组合(长2000;皮特森2000;皮特森et al . 2003年)。过多的养分输入河口层叠效果,减少底栖生物多样性(豪沃思et al . 2000年),并增加有害藻华的频率和缺氧或缺氧(伯施et al . 2001年)。减少的植被(豪沃思et al . 2000年)和简化的群落结构由于增加缺氧事件的频率和持久性(迪亚兹和罗森博格1995)。
从海水养殖废水可能导致富营养化和当地沉积的变化,所有这些影响底栖生物多样性和底栖生物的功能作用在半封闭的生态系统如河口、海湾、和峡湾(Naylor et al . 2000年)。很少有和有限的海水养殖废水的效果测量水柱(麦金农et al . 2002年),和大多数影响沉积物中看到(Ervik et al . 1997年)。沉积过剩的食物残渣和粪便mari-culture附近的设施导致局部低氧/缺氧和底栖生物多样性的减少。只要一个影响区域不是已经从其他来源over-enriched,额外的营养负荷预计不会有实质性的大规模对ben-thic生物多样性的影响。饲养在海水养殖生物滤食甚至可能改善水质和富营养化抵消负面影响底栖生物多样性。适当大小的旨在减少栖息地的破坏和海水养殖操作,关注生产的本地物种低食物网(双壳类和食草鱼类),可能对底栖生物多样性构成什么威胁(Naylor et al . 2000年)。
河口之间的大陆架和通过接口和货架的水域,污染物的威胁还包括直接和间接的输入(克拉克1997年)。一些污染物排放通过管道直接放到架子上或从船只倾倒。间接通过排入河流和河口输入发生,并通过空气的污染雨水。缺氧可以在小的压力和增加死亡率和大型生物(灰色et al . 2002年),和在较大的生物如鱼(迪亚兹和罗森伯格1995)。由此产生的微生物群落和生产中断影响沉积物中污染物的氧化过程和解毒。倾倒垃圾在更深的水域货架和溢出的石油设施和船只也导致污染远离沿海的利润率。沉船和碰撞导致大规模污染(彼得森2000;彼得森et al . 2003年)使用效果可以持续多年(例如,超过10年Dauvin 1998),尽管泄漏在开放海域的空间尺度上往往是相对于栖息地面积小,和累积的影响是未知的。
倾销疏浚材料导致栖息地的改变通过窒息和toxic-ity(索莫菲尔德et al . 1995年)。变更的水流入河流和疏浚河道的水文变化书架,盐度、泥沙沉积(·德容&·德容2002)。影响可能更严重比河口因为大陆架生物生理上似乎不那么宽容这些类型的干扰。
材料倾倒在深海包括常规弹药和化学武器(施里弗et al . 1997),低收入和中级放射性废物(泰尔2003),污水污泥(Bothner et al . 1994),疏浚破坏包含污染物(泰勒2003年),和各种船只和结构与军事有关,运输和石油和天然气开采(施里弗et al . 1997年)。现代深海研究显示,这个生态系统动态和活性比此前认为的泰勒(2003)和在很大程度上,西方国家停止深海倾销。雷竞技手机版app深海现在被视为多余的二氧化碳存储库(赫尔佐格et al . 2000年),这是液体在高压力和低温度(史密斯Glover & 2003)。深海二氧化碳处理可以降低大气中的二氧化碳增加和减少海洋表面的pH值,但同时降低深海pH值(卡尔德拉&达菲2000)致命水平的二氧化碳羽流(史密斯Glover & 2003)。
大陆斜坡表现出最高的碳沉积,也许最大的动物生产力和生物多样性最高的深海(雷克斯1983)。因为他们也最容易深海环境中,人类活动可以有更大的对生物多样性的影响和关键斜坡沉积过程等碳埋藏或生产。很少有研究解决这些问题的深海。
深海浓缩实验(Snelgrove et al . 1992年)表明,有机的输入选择短寿,surface-dwelling,投机取巧的物种,和类似的效果可能发生处置污水污泥和疏浚材料在深海。浓缩也可以改变饮食和局部加强浅海底物种(Grassle 1991;范多佛et al . 1992年)。物理干扰从底拖网也产生机会主义,缺乏多样性组合。更换更大,deeper-dwelling物种小投机者降低生物扰动作用,限制了沉积物的氧化,减缓碳降解和埋葬。斜率大型生物一般消费和埋葬(10厘米或更多)新鲜有机质在几天内到达海底(伯爵1989;莱文et al . 1997年),所以最不稳定的碳是气息奄奄的迅速或掩埋。在深海深处,可能发生人为抑制生物扰动作用和碳埋葬二氧化碳封存或结节采矿活动,效果类似于自然质量下滑(马森et al . 1994年)。
大规模的威胁(盆地规模):气候变化raybet雷竞技最新
raybet雷竞技最新气候变化将改变盐度、温度和风的模式,这将会影响当地的、地区和大规模水文学(Manabe et al . 1994),但这可能发生缓慢。例如,海平面上升将在架子上或很少有直接影响深海系统,但它会产生实质性的影响沿海环境(史密斯et al . 2000年)。同样,海平面上升造成损失的滩涂和湿地向陆地迁移(Donnelly & Bertness 2001),但海岸线铠装都限制了这一运动,导致潮间带河口栖息地的丧失(Pethick 2001)。因为扎根植被是一个关键部分河口湿地生境结构和初级生产的大规模损失与海平面上升(彭et al . 2003年)构成严重威胁生物多样性和生态系统过程和服务(莫里斯et al . 2002年)。
在更大的空间尺度上,影响可能会导致频率的变化防御气象现象如ENSO和海洋(厄尔尼诺现象)NAO(北大西洋涛动)事件和边界强度的电流。
raybet雷竞技最新气候变化改变季节性的降雨模式对深海系统或温度几乎没有影响。然而,在沿海地区,气候变化可能会导致当地物种灭绝的流行底栖raybet雷竞技最新无脊椎动物适应环境变化的历史模式,同时促进一些入侵物种的范围扩张。尽管许多入侵物种在河口广盐性的(能够容忍各种盐度),入侵发生在高的比低盐度(Ruiz et al . 2000年)。减少淡水流允许穿透深入河口海洋水域,并成功的介绍,其中包括病菌(如寄生腰鞭毛虫,Hematodinium spp),期间更频繁干旱或减少淡水流入系统(卡尔顿et al . 1990;梅西克et al . 1999年)。有明显差异很大河口底栖生物群落的抗入侵(Ruiz et al . 2000),硬衬底社区和证据表明,多样性有助于抵抗入侵(Stachowicz et al . 2002年)。与气候变化,因此合理的期待转变入侵阻力和丰富的入侵者。
raybet雷竞技最新气候变化是增加极端天气的频率和规模,和强风增加波浪作用,导致物理干扰到海底浅大陆架海域海岸线和霍尔(1994)。效果类似于广泛的渔业干扰。在浅地区,风致波浪作用实际上可能增加养分循环由于沉积物的物理干扰,提高污染物通过增加沉积物的氧化解毒。在正常情况下,架子上底栖生物在秘鲁的智利上升流区往往氧气压力,较低的生物多样性、生物量低,和小生物扰动作用的活动。厄尔尼诺上升流代替通常用氧化水缺氧大陆架水域质量,促进更大的底栖生物多样性、生产力、养分循环和粮食生产。这类型的气候raybet雷竞技最新事件可能提高大多数海洋货物和服务(Tara-zona et al . 1988;古铁雷斯et al . 2000年)。
附加在夏季冬季降雨量和降雨在温带沿海水文特征和变化可能影响沉积物加载和洪水和侵蚀控制的有效性由造礁和sediment-stabilizing生物。加速冰川融化,大规模减少盐度(其中一些将抵消增加筑坝河流),和增加的温度会改变底栖生物群落结构和多样性(史密斯et al . 2000;奥斯丁et al . 2002年)。温度可提高生产率的提高,营养不限制,导致积极的影响的一些海洋底栖生物提供的商品和服务。
对潜在影响紫外线辐射增加海洋沉积底栖生物,但从事远洋鸡蛋表明鱼类(Kouwenberg et al . 1999年)表明,影响浮游幼虫阶段是可能的。
系统之间的比较
我们总结了脆弱性不同威胁生态系统的三个分组(表7. a1-7。A3附录184页),当他们与供应的商品(如食物、纤维)和服务(例如,水过滤、防洪、废物回收),以及支持生态系统进程(例如,固碳、养分循环、分解),栖息地的维护服务,和审美服务(精神浓缩,娱乐,科学探究)。因为许多可用的数据漏洞轶事或收集和解释在非常不同的框架不同的威胁,客观、定量比较跨系统是不可能的。我们使用我们的集体经验和现有研究生态系统过程和服务开发一个定性等级方案来解决不同系统的相对脆弱的服务或流程(表7. a1-7。A3附录184页)。在某些情况下,一个给定的威胁可能会提高某些服务或流程,因此负分数是可能的。例如,引入物种可能创建生态破坏,但如果它是可食用的和丰富的,它可能会增加供应的食物。
我们从这个练习,河口系统解释目前我们三个大致分类的最脆弱的海洋系统,部分是因为广泛的服务进行的环境和部分原因是强度和数量的威胁。远程深海系统最脆弱的海洋系统根据现行模式的剥削,因为他们的大区域互联的栖息地和暴露于人类活动相对较低。因为它的大小,大到中档效应(如气候变化)的最关心的问题。raybet雷竞技最新尽管如此,代表一个地区的极端关注海山,和大陆斜坡易受人类开发的局部效应的增加。
恢复,恢复和恢复海洋生态系统
海洋恢复陆地和淡水生态系统的落后,部分原因是海洋是巨大的规模和共同所有权,这阻碍了干预。近年来,然而,一个框架,用于恢复,恢复和康复(见Frid &克拉克1999和霍金斯et al . 1999定义)海洋生态系统的发展,特别是在沿海系统由habitat-providing生物群如海草床,红树林,盐沼(Ewel et al . 2001年)。
在开放海洋生态系统中,幼虫传播障碍很少,而且有很好的潜力自然恢复unimpacted开拓殖民地的人口。可以迅速而自然恢复,积极恢复在开放海洋系统是困难的除外生物结构创建并维护的栖息地。例如,生物生成的结构如海草、盐沼泽盐生植物,和红树林可能分散不好(奥尔特et al . 1994),但积极的种植可以加速栖息地恢复和生态系统过程和相关服务。
封闭的水域,如河口、海湾和泻湖可以经得起修复水质和海洋冲洗的操作。例如,大型植物可以吸收的营养物质在半封闭的地区,可能成为富营养的。Phy-toplankton现存量也可能受到滤食动物(官et al . 1982;戴维斯et al . 1989;已有1991人)。大多数海洋系统的开放性意味着水质可以提高区域减少有害的输入和活动。海洋潮汐冲洗的限制,例如,通过沉降道路建设或情景入口关闭,栖息地可以恢复活跃的疏浚,操作流程,并建立永久开放水湾。
许多近岸和货架地区严重影响干扰效果可以减轻只有大面积安静的离开。提出了海洋保护区作为务实的防范全球渔业管理工具与更广泛的海洋保护的好处(2004年美国海洋政策委员会)。这些可以有各种级别的保护(见詹宁斯&凯泽1998和大厅1999优秀概述)从绝对排斥(“不带区”)那么严格地区齿轮类型是有限的或钓鱼是排除在某些季节。
大多数沿海修复工作都集中在特定的栖息地,生物栖地、组合、或物种,但沿海生态系统强烈相互联系和协调的努力有时是必要的。因此,海草恢复可能援助盐泽恢复或修复。积极恢复有最强的层叠效果,”生态系统工程师”(劳顿1994)。牡蛎礁、河蚌、海草床、盐沼和红树林都所赋予的例子,一个强大的结构元素是占主导地位的生物群。的大型植物(如海草)可以种植形成核的新床(丰塞卡et al . 2002年)。相反,牡蛎礁修复不成功的盐沼和海草床附近,提供走廊大型移动捕食者(Micheli &彼得森1999)。
结论
我们的讨论集中在海洋沉积物的许多流程和服务,在某些情况下现在才被认可。关键栖息地的重要性,比如利用少年鱼类,已成为一个主要关注渔业组织仅在过去十年左右的时间。有未来的服务和流程,不断恶化,但接收没有注意,因为他们还不承认吗?这是未知的未来价值的海洋沉积系统,将一个特定的紧迫性保留剩下的仍相对原始的系统;服务和流程的潜在损失,我们这里有总结可能只是故事的一部分。最终,生物多样性本身有价值,超越产品和服务提供给人类和生物多样性的流程可以帮助支持。但即使是对于那些未能识别的美丽生物的多样性和道德义务保护这种多样性,潜在的损失的货物和服务支持的海洋沉积动物群至少应该提供暂停(和提供食物)认为,保护这些生物资源和动力。
文献引用
Ailstock,硕士:诺曼,P.J. Bushmann》2001。常见的芦苇芦苇aus-tralis:控制和影响淡水nontidal湿地生物多样性。恢复生态学9:49-59。
奥斯丁,m。,PJ.D。Lambshead,我!钦斯·g·鲍彻,c .敏感性g .国王,小池百合子,C.R.史密斯,P.V.R. Snelgrove》2002。生物多样性的海洋湿地之上和之下的链接,可能会影响社会稳定。生物多样性和保护11:113 - 136。
南风,P.J.,以R.W.兰顿R.J. Malatesta l·沃特电子商务。情人节,C.L.S.唐纳森,E.W.兰顿,A.N. Shepard, I.G.巴伯。1996。移动渔具的影响在墨西哥湾海底栖息地缅因州(西北大西洋):对保护鱼类种群的影响。渔业科学评论4:185 - 202。奥斯特PJ。、R.J. Malatesta和南卡罗来纳州LaRosa。1995年。小环境利用的模式移动巨型动物新英格兰(美国)南部大陆架和斜率。《海洋生态发展系列127:77 - 85。 Ayres, D.R., D. Garcia-Rossi, H.G. Davis, and D.R. Strong. 1999. Extent and degree of hybridization between exotic (Spartina alterniflora) and native (S. foliosa) cordgrass (Poaceae) in California, USA determined by random amplified polymorphic DNA (RAPDs). Molecular Ecology 8:1179-1186. Baker, K., and R.L. Haedrich. 2003. Could some deep-sea fishes be species-at-risk? International Deep-Sea Biology Conference, Aug. 2003, Coos Bay, Oregon (Abstract).
彭,J。美国Beville, d . Britsch哈特利,s·霍斯,j·约翰逊·坎普Kinler, a . Martucci j . Porthouse d·里德k·罗伊,s .萨普克塔和j . Suhayda》2003。历史和预测1978 - 2050年路易斯安那州沿海土地变化。03334年美国地质调查局的打开的文件报告,36页。
布鲁姆,H。g·施里弗,h·泰尔》1995。Megabenthic实验干扰深海锰结核地区开拓殖民地。海洋Georesources和印度尼西亚13:393 - 416。
伯施,测向、相对湿度Burroughs J.E.贝克,R。P梅森,中一段罗,R.L.谢菲特。2001。海洋污染在美国。阿灵顿,弗吉尼亚州:皮尤海洋委员会。
博罗夫斯基,c . 2001。身体受到干扰,在秘鲁盆地深海大型生物,东南太平洋重新审视后7年实验的影响。深海研究II 48:3809 - 3839。
Bothner, M.H.高田h,骑士,r·希尔,b . Butman J.W.法灵顿,水银血压计Colwell和参考书籍Grassle。1994。污水污染沉积物下深海垃圾场了纽约。海洋环境研究38:43-59。
Butman, c.a、j.t卡尔顿和共和国所有。1995。捕鲸对深海生物多样性的影响。保护生物学9:462 - 464。
卡尔代罗,K。2000年,P.B.达菲。。南大洋的角色人为二氧化碳的吸收和存储。科学287:620 - 622。
卡尔顿,j.t 2001。介绍了物种在美国沿海水域:环境影响和管理重点。阿灵顿,弗吉尼亚州:皮尤海洋委员会。
卡尔顿,j.t和j·b·盖勒。1993。生态轮盘:全球运输非土著海洋生物。科学261:78 - 82。
卡尔顿,j.tj•k•汤普森,担忧也名列前茅,的L.E.·舒梅尔和F.H.尼克尔斯》1990。非凡的入侵旧金山湾(美国加州)由亚洲蛤Potamocorbula amurensis介绍和传播。《海洋生态发展系列66:81 - 94。
查宾,F.S.三世,P.A. Matson, H.A.穆尼。2002。陆地生态系统生态学的原则。纽约,气象出版社。
克拉克,1995年核磁共振。罗非鱼的经验与管理(Hoplostethus atlanti-cus)在新西兰水域,股票和商业捕捞的影响在此期间1980 - 1993。:深海渔业的北大西洋海洋斜率,编辑A.G.料斗,251 - 266页。多德雷赫特,荷兰,Kluwer学术出版商。
克拉克,R.B.,编辑器。1997年。海洋污染(第四版)。牛津,克拉伦登出版社。
科恩,清醒,右眼Giotta、M.W. Luckenbach D.L. Breitburg》1999。牡蛎礁功能、增强和恢复:栖息地的商业化开发和利用——和重要生态物种。18:712贝类研究》杂志上。
牧羊犬,j.s中华民国大厅,M.J.凯撒,起始点效果。2000。货架海钓的底栖生物扰动:趋势和预测。动物生态学杂志69:785 - 798。
骗子,正当,苏尔。1999。在人口爆炸的入侵物种:滞后次原因和影响。:外来物种入侵和生物多样性管理、ot Sandlund编辑,P.J. Schei, a .威肯,103 - 126页。波士顿,Kluwer学术出版商。
Dauvin J.C. 1998。湾的细沙磨料alba社区粗亚麻布阿莫科加的斯石油泄漏后二十年。海洋污染公告36:669 - 676。
戴维斯,开国元勋之一B.R.安贝德卡对、诉斯图尔特和m . de Villiers》1989。过滤serpulid保利的活动
chaete人口(Ficopomatus enigmaticus Fauvel)及其对水质的影响在沿海码头。河口海岸和大陆架科学29:613 - 620。
戴维斯。、洛杉矶莱文和沃尔特。2002。人工装甲海岸线:开放沿海社区引入到加州南部海湾。海洋生物学140:1249 - 1262。
P.K.代顿市1972年,水银血压计海瑟。。生物扰动作用在维持多样性的深海。深海研究19:199 - 208。
P.K.代顿市旧金山那里画眉,M.T. Agardy, r Hofman。1995。环境影响海洋渔业。水生保护:海洋和淡水生态系统5:205 - 232。
P.K.代顿市,美国画眉,科尔曼。2002。渔业海洋生态系统的生态影响美国。弗吉尼亚州阿灵顿,皮尤海洋委员会。
Deegan,洛杉矶J.W.天,Jr .) J.G. Gosselink, a . Yanez-Arancibia g . Soberon查韦斯,p . Sanchez-Gil。1986年。物理特性之间的关系,植被分布并在墨西哥海湾河口渔业产量。:河口可变性,由沃尔夫D.A.编辑,页83 - 100。奥兰多,佛罗里达,学术出版社有限公司
·德容,V.N.2002年,D.J.·德容。。“全球变化”的影响在排水年际变化驱动因素在河里疏浚和废渣处理莱茵系统和浊度的瓦登海。河口海岸科学55:969 - 991和货架。
Diaz R.J.,r·罗森博格。1995。海洋底栖生物缺氧:回顾其生态效应和底栖大型生物的行为反应。海洋学和海洋生物学:年度审查33:245 - 303。
唐纳利,J。P,医学博士Bertness》2001。快速向岸侵蚀的盐沼带状草地在回应加速海平面上升。美国国家科学院院刊》上,美国98:14218 - 14223。
达菲,J.E.,干草。2000。强烈影响放牧片的底栖生物群落的组织。生态专著70:237 - 263。
Ervik,。,P。K. Hansen, J. Aure, A. Stigebrandt, I! Johannessen, and T. Jahnsen. 1997. Regulating the local environmental impact of intensive marine鱼类养殖,妈妈的概念系统(Modelling-Ongrowing鱼farms-Monitoring)。水产养殖158:85 - 94。
Ewel, kc,c . Cressa R.T. Kneib,注:湖,洛杉矶Levin硕士帕默,p . Snelgrove d.h。2001。管理关键的过渡区。生态系统4:452 - 460。
Folke C。美国木匠,t·姆l·甘德森C.S.温和和沃克。2002。弹性和可持续发展:建立适应能力的世界里转换。中31:437 - 440。
塞卡M。,情感表达。凯利·维特菲尔德,新墨西哥州,贝尔和S.S.。2002。建模海草景观格局和生态属性相关联。生态学应用12:218 - 237。
Frid, C.L.J.克拉克,美国。1999年。恢复水生生态系统:概述。水生保护:9:1-4海洋和淡水生态系统。
格洛弗,A.G.,C.R.史密斯。2003。深海层生态系统:现状和前景人为改变到2025年。环境保护30:1-23。
Goldburg,效力位于美国罗德岛州,硕士艾略特,Naylor》2001。海洋水产养殖在美国:环境影响和政策选择。弗吉尼亚州阿灵顿,皮尤海洋委员会。
伯爵,g . 1989。Benthic-pelagic耦合在深海底栖生物群落。自然341:437 - 439。
1991年Grassle,参考书籍。污水污泥对深海的影响社区(抽象)。EOS、事务的美国地球物理联盟72:84。
Grassle,参考书籍,新泽西州Maciolek》1992。深海物种丰富度:地区和地方多样性估计从定量下样品。美国博物学家139:313 - 341。
Grassle,参考书籍1973年,上半叶桑德斯。。生命的历史和扰动的作用。深海研究20:643 - 659。
灰色,j.s、吴R.S.S.陈文贤或。2002。缺氧的影响和有机浓缩在沿海海洋环境。《海洋生态发展系列238:249 - 279。
古铁雷斯,D。退役军人盖拉多,美国市长,c . Neira, c . Vasquez j . Sellanes m·里瓦斯a·索托f·卡拉斯科,m·巴尔。2000。溶解氧的影响和新鲜有机质在大型生物的生物扰动作用潜力潮下带沉积了智利中部在1997/1998的厄尔尼诺现象。《海洋生态发展系列202:81 - 99。
大厅,s . 1994。物理干扰和海洋底栖生物群落:生活在疏松的沉积物。海洋学和海洋生物学:年度审查32:179 - 239。
大厅,s . 1999。钓鱼对海洋生态系统的影响和社区。牛津布莱克韦尔科学。
Hallegraeff,通用1992年,C.J. Bolch。。运输的硅藻和腰鞭毛虫休眠孢子在船舶压载水:对浮游生物的生物地理学和水产养殖的影响。《浮游生物研究14:1067 - 1084。
Hall-Spencer, J。、V艾蓝和J.H.窝》2002。拖网捕鱼破坏东北大西洋古老的珊瑚礁。伦敦系列的《皇家学会学报B 269:507 - 511。
Hall-Spencer, J.M.,打开摩尔。2000。扇贝疏浚具有深远的长期影响maerl栖息地。冰海洋科学杂志57:1407 - 1415。
霍金斯,中华民国、jr艾伦和布雷。1999。温带海洋和沿海生态系统的恢复:逼近性质。水生保护:9:23-46海洋和淡水生态系统。
赫尔佐格,H。b . Eliasson, o . Kaarstad》2000。捕获温室气体。科学美国人282:72 - 79。
沪元,c . 1991。是底栖生物的活动暂停喂一个因素控制在布雷斯特湾的水质?《海洋生态发展系列69:179 - 188。
钟,硕士,P。D. Boersma, M.L. Hunter, Jr., F. Micheli, E.A. Norse, H.P Possingham, and P.VR. Snelgrove. 2001. Oceans at risk: Research priorities in marine conservation biology. In: Conservation Biology: Research Priorities for the Next Decade, edited by M.E. Soule, and G.H. Orians, pp. 125-154. Washington, DC, Island Press.
豪沃思,R。安德森,j . Cloern c . Elfring c .霍普金森LaPointe, t·马龙,n . Marcus k . McGlathery a . Sharpley和d·沃克。2000。营养沿海污染河流、海湾和海洋。生态学7:1-15问题。
杰克逊,J.B.C.,w•h•伯杰M.X. Kirby·Bjorndal, l博茨,,B.J. Bourque r.h布拉德伯里,r·库克j . Erlandson J.A.埃斯蒂斯,t休斯,s . Kidwell C.B.兰格,H.S. Lenihan, J.M. Pandolfi, h彼得森,R.S. Steneck M.J. Tegner,水银血压计华纳。2001。历史过度捕捞和最近的沿海生态系统的崩溃。科学293:629 - 638。
Jassby,公元j•Kimmerer,密度Monismith, c .装甲J.W. Cloern, T.M.鲍威尔,jr Schubel, T.J. Vendlinski》1995。Isohaline位置作为河口生境指标数量。生态学应用5:272 - 289。
詹宁斯。,M.J.凯撒。1998。钓鱼对海洋生态系统的影响。在-352年海洋生物学34:201进步。
Kaiser M.J.k·拉姆齐c.a理查森,远东斯宾塞,境品牌。2000年。慢性钓鱼扰动改变了架子上海洋底栖生物群落结构。动物生态学杂志69:494 - 503。
凯撒,M.J.1994年,B.E.斯宾塞。。鱼清除最近在地区的行为。《海洋生态发展系列112:41-49。
Kalke,进食和爸爸。Montagna》1991。在大型底栖生物的影响淡水流入Lavaca河三角洲和上Lavaca湾,德克萨斯州。在海洋科学贡献32:49 - 71。
汗,R.A. 1987。影响慢性接触石油碳氢化合物在两种海洋鱼类感染hemoprotozoan,锥虫属murmanensis。加拿大《动物学65:2703 - 2709。
Kneib,保留时间1991。间接影响的实验研究海洋柔软的沉淀物的社区。美国zoologist31:874 - 885。
Kneib,保留时间2000。河口盐沼全海洋和生产转移的自游生物在美国东南部。:在海涂生态概念和争议,由议员编辑温斯坦和D.A.克雷格,267 - 292页。多德雷赫特;波士顿,Kluwer学术出版商。
Koslow,正当,J.D.戈登G.W. Boehlert R.L. Haedrich, p . Lorance, n .标准杆。2000。大陆坡和深海渔业:对脆弱生态系统的影响。冰海洋科学杂志57:548 - 557。
Koslow,正当k .稀有J.K. Lowry, t·奥哈拉G.C.B.波尔,a·威廉姆斯。2001。海底山底栖大型生物塔斯马尼亚南部:社区的结构和影响拖网捕鱼。《海洋生态发展系列213:111 - 125。雷竞技csgo
Kouwenberg, J.H.M.做艾滋病病毒,Browman J.J.卡伦,水戴维斯,肯尼迪。圣皮埃尔,J.A.龙格。1999。生物权重的紫外线(280 - 400 nm)引起的死亡率在海洋浮游动物和鱼。即大西洋鳕鱼(Gadus morhua)蛋。海洋生物学134:269 - 284。
Lambshead PJ.D。2003,g .鲍彻。。海洋线虫深海diversity-hyper-diverse还是炒作?生物地理学杂志30:475 - 485。
兰斯洛特,C。,j . Staneva d . Van Eeckhout j·贝克斯,大肠Stanev。2002。造型Danube-influenced西北大陆架的黑海。二:生态系统响应的变化营养交付后由多瑙河1972年筑坝。河口、沿海和货架科学54:473 - 499。
劳顿,J.H. 1994。在生态系统的物种做什么?Oikos 71:367 - 374。
勒尼汉,H.S.1998年,h彼得森。。如何通过渔业干扰生境退化提高缺氧对牡蛎珊瑚礁的影响。生态学应用8:128 - 140。
Lennsen J.EM。,F.B.J.表示“状态”,w•h•范德Putten C.W.P.M.布鲁姆》2000。物种组成的变化和物种丰富度在芦苇南极光河岸区域主导。植物生态学147:137 - 146。
Lerberg, S.B.、自动跟踪荷兰和桑格。2000。反应的潮沟macroben-thic社区流域发展的影响。河口23:838 - 853。
莱文,洛杉矶:布莱尔,g . Plaia D.J. DeMaster w·福尔马丁c和c·托马斯。1997。有机物的快速俯冲Maldanidpolychaetes在北卡罗莱纳的斜率。ofMarine研究杂志55:595 - 611。
莱文,洛杉矶怀特测向,a . Covich c·达姆c . Erseus K.C. Ewel R.T. Kneib, a . Moldenke硕士帕默,p . Snelgrove d不走正路,J.M. Weslawski。2001。海洋的功能关键的过渡区和沉积物生物多样性的重要性。生态系统4:430 - 451。
刘易斯,剩下凯利,j·达文波特,郭台强。2002。管道施工的影响的研究河口底栖无脊椎动物社区。河口、沿海和货架科学55:213 - 221。
长,急诊室》2000。美国河口沉积物质量退化:回顾大小和生态意义。生态学应用10:338 - 349。
麦金托什,这位,提到过阿什顿和美国Havanon》2002。红树林恢复和潮间带生物多样性:在拉廊红树林生态系统的一项研究中,泰国。河口、沿海和货架科学55:331 - 345。
Manabe, S。、R.J.史都华牌和M.J.斯佩尔曼。1994。反应耦合的海洋大气模型增加大气中的二氧化碳。中23:44-49。
马森,D.G.ibsen Pinheiro, J.A.卡特赖特,L.M. R.B.惠特马什,他们Beslier规则,h·罗斯。1994。压缩变形在东北大西洋ocean-continent过渡。伦敦地质学会的期刊151:607 - 613。
麦金农,公元Trott洛杉矶,m . Cappo位米勒,达根,p . Speare, a·戴维森。2002。营养的命运虾的农场污水在红树林小溪北昆士兰,澳大利亚。河口、沿海和货架科学55:655 - 671。
梅里特,天然橡胶1997年,R.L. Haedrich。。深海底栖鱼类和渔业。伦敦,查普曼和大厅。
Messick G.A.中华民国约旦和W.F. van Heukelem》1999。盐度和温度影响Hematodinium sp.的蓝蟹Callinectes sapidus。《贝类研究18:657 - 662。
Micheli F。1999年,h彼得森。。河口栖息地植被作为捕食者运动的走廊。保护生物学13:869 - 881。
矿产资源管理服务》2004。2004年墨西哥湾深水:美国的边界扩张。美国内政部矿产管理服务,商务地区墨西哥湾。商务报告MMS 2004 - 021。
摩尔,M.J.智慧化Smolowitz, J.J.斯蒂。1997。阶段的浮肿的空泡形成的肝脏冬季比目鱼Pleuronectes也从化学污染。水生生物的疾病31:19-28。
莫里斯,j.t,P。b . Kjerfve V Sundarweshwar,比较温度Nietch,湄Cahoon。2002。的反应沿海湿地海平面上升。生态83:2869 - 2877。
迈尔斯,R.A.,蠕虫。2003。全球快速损耗的食肉鱼类的社区。自然423:280 - 283。
国家研究委员会,1995年。了解海洋生物多样性。华盛顿特区国家科学院出版社。
那,R.L.、R.J. Goldburg J.H.白桃花心木:考茨基,M.C.M.贝弗里奇,j .粘土c . Folke j .卢布琴科h·穆尼和m . Troell。2000。水产养殖对世界鱼类供应的影响。自然405:1017 - 1024。
纽厄尔司令部l·j·Seiderer,湄希区柯克。1998。在沿海水域疏浚工程的影响:回顾对扰动的敏感性和随后的复苏的海底生物资源。海洋学和海洋生物学:年度审查36:127 - 178。
官C.B.,T.J. Smayda, r·曼。1982。底栖生物过滤器喂养:自然eutroph-ication控制。《海洋生态发展系列9:203 - 210。
Orensanz, J.M.,连、d·阿姆斯特朗和r . Hilborn》1998。甲壳类动物资源很容易串行depletion-the多方面的螃蟹和虾渔业的下降更大的阿拉斯加湾。评论在鱼类生物学和鱼类8:117 - 176。
奥尔特,效力、m . Luckenbach·摩尔》1994。在海洋大型植物传播种子:对殖民和恢复的影响。生态75:1927 - 1939。
Ozturgut E。J.W. Lavelle,右眼烧伤。1981。锰结核开采对环境的影响:在北赤道太平洋半工业规模开采测试结果。:海洋环境污染2:倾销和采矿,由美国编辑盖尔,页437 - 474。爱思唯尔海洋学系列。阿姆斯特丹,爱思唯尔。
2002年共和国所有。海洋保护区:生态系统管理和保护的工具。弗吉尼亚州阿灵顿,皮尤海洋委员会。
保利,D。克里斯滕森,V, j . Dalsgaard r .森林,f·托雷斯。1998。海洋食物网捕鱼了。科学279:860 - 863。
保利,D。和j·麦克莱恩。2003年。在一个完美的海洋:国家渔业和海洋生态系统在北大西洋。华盛顿特区岛出版社。
彼得森,2000 h。在阿拉斯加埃克森·瓦尔迪兹号漏油事件:急性、间接和慢性对生态系统的影响。海洋生物学的进步39:3 - 84。
彼得森,h美国南达科他州,大米,J.W.短,d . esl J.L.锥子B.E. Ballachey、抓铁。2003。长期生态系统应对埃克森·瓦尔迪兹号漏油事件。科学302:2082 - 2086。
Pethick, j . 2001。沿海管理和海平面上升。系列42:307 - 322。
拉姆齐K。,M.J.凯撒。1998。水底的渔业干扰增加海螺捕食风险(Buccinum undatum l .)。《海洋研究39:299 - 304。
拉姆齐K。,M.J.凯撒,雷诺数休斯。1998。底栖生物食腐动物捕鱼扰动的响应拖齿轮在不同的栖息地。实验海洋生物学和生态学杂志224:73 - 89。
雷克斯,1983年硕士。物种多样性的地理模式深海海底。:大海。深海生物,编辑G.T.罗,页453 - 472。纽约约翰·威利和儿子。
Roehner, M。r . Bastrop附近,k . Juerss》1996。欧洲的殖民两名美国遗传类型或种类的属Marenzelleria(多毛纲:海稚虫科):异型酶的电泳分析。海洋生物学127:277 - 287。
罗斯柴尔德,B.J.,j.s奥尔特,p . Goulletquer和m . Heral。1994年。下降的切萨皮克湾牡蛎人口:一个世纪的栖息地的破坏和过度捕捞。《海洋生态发展系列111:29-39。雷竞技csgo
鲁伊斯,通用,P。W. Fofonoff, J.T. Carlton, M.J. Wonham, and A.H. Hines. 2000. Invasion of coastal marine communities in North America: Apparent patterns, processes and biases. Annual Review of Ecology and Systematics 31:481-531.
Salen-Picard C。,2002 d Arlhac。。在地中海底栖生物群落长期变化:多毛纲的组合和水文变化之间的关系的罗纳河。河口25:1121 - 1130。
施里弗,G。a . Ahnert h·布鲁姆,c .博罗夫斯基,h·泰尔》1997。大型深海环境影响研究的结果远程协作学习在八年的调查。学报》第七届(1997)国际海洋和极地工程会议,1:438 - 444。
Sklar, F.H.1998年,J.A.布劳德。。沿海变化所带来的环境影响淡水流入墨西哥湾。环境管理22:547 - 562。
史密斯C.R.贝拉·奥斯丁·g·鲍彻,c .敏感性g·王,小池百合子,p·哈钦斯j . Lambshead, PVR。Snelgrove》2000。全球变化和生物多样性的联系在湿地中。生物科学50:1076 - 1088。
Snelgrove, PVR。、参考书籍Grassle和水Petrecca》1992。在保持较高的深海食物补丁的作用多样性:田间试验与水动力地无偏殖民托盘。湖沼学和海洋学37:1543 - 1550。
索莫菲尔德,P.J.上半叶里斯和m .沃里克。1995。相互关系在浅水海洋较小型底栖生物群落结构和大型生物与疏浚处置。《海洋生态发展系列127:103 - 112。继续阅读:微片栖息地生态景观区域
这篇文章有用吗?

