研究地点和方法
研究网站包括centreeast 13森林,东南,南海岸,西南地区,和科特迪瓦的西南。这些不同的森林的选择是基于完整无缺的森林覆盖和物种列表的可用性。从半落叶到常绿的森林(图5.2),它们是各种大小(2590 - 300000公顷),并有不同的土壤和气候条件(表5.1)。三种土壤类型可以区分:三级砂、粘土土壤来源于片岩,或沙土造成花岗岩风化(De Rham 1971)。总年降雨量范围在1400 - 2300毫米,而水分亏缺是150至400毫米。干燥季节的长度变化2至5个月(Perraud 1971)。
本研究中使用的大多数的数据来自不同的研究,在清单的森林(表5.1)。这样的库存目标识别所有的植物类群在森林里遇到而遵循已有的路径或路径创建。此外,我们进行库存那些没有或森林
地理区域 |
森林 |
的缩写 |
森林类型1 |
面积(公顷) |
坐标 |
年降雨量”(毫米/年) |
东方中心 |
Bossematie |
Bm |
SD |
22200年 |
6°20 ' - 6°35镑3°20 - 3°35 'w |
1400 - 1500 |
东南 |
Mabi |
Mb |
E |
59616年 |
5°51 - 6°05’3°22 - 3°4 l 'W |
1650 - 1700 |
Songan |
年代 |
E |
38189年 |
5°46 - 6°12镑3°12 - 3°26 'w |
1600 - 1650 |
|
Tamin |
T |
E |
24934年 |
5°40 - 5°58镑3°11的25 'w 3° |
1650 - 1700 |
|
亚亚 |
Y |
E |
23877年 |
5°35 - 5°54他3°23 - 3°46 'w |
1700 - 1800 |
|
南海岸 |
Dassioko |
D |
E |
11203年 |
5°00 - 5°07年5°50镑的- 5°57 'w |
1550 - 1600 |
Monog ^ ga |
莫 |
E |
39660年 |
4°44 ' - 4°58镑6°14 - 6°35 'w |
1650 - 1750 |
|
港口附近 |
PG |
E |
2590年 |
5°06 - 5°11镑5°29 - 5°34 'w |
1550 - 1600 |
|
银行 |
B |
E |
3300年 |
5°21 - 5°25镑4°01的4°05 'w |
2000年 |
|
西南地区 |
上流社会的Sassandra |
海关 |
SD |
102400年 |
6°22 - 7°24’6°59的7°10 'w |
1460 - 1680 |
Marahoue |
马 |
SD |
101000年 |
6°53 - 7°14镑5°46的10 - 6°'w |
1400年 |
|
西南 |
高级渡渡鸟 |
高清 |
E |
236733年 |
4°41 - 5°19他7°01”- 7°25 'w |
1900 - 2300 |
大 |
大 |
E |
300000年 |
5°09年“啊——6°09年6°48”- 7°26 'w |
1800 - 2200 |
1。盖伊表示:&Adjanohoun (1969)
2。Eldin (1971)
3所示。De Rham Perraud & Souchere (1970), (1971)
下层土壤和土壤 |
水deficit4(毫米/年) |
分类单元(#) |
源 |
片岩 |
350 - 400 |
611年 |
Ake Assi (1992) Bakayoko (1999) |
片岩 |
300年 |
640年 |
本研究 |
片岩 |
300年 |
591年 |
本研究 |
片岩 |
200年 |
512年 |
本研究 |
本研究Schis: 200 617
沙450 719 Ake Assi (1997)
沙300 859 Kouame (1998 b)
沙450 705 Kouame (1998 b)
沙200 773 De今敏(1983)
花岗岩400 843 Kouame (1998)
片岩350 - 400 475 Jongkind等人工智能(1999)
花岗岩150 906 Kouadio (2000)
Kouassi (2000)
花岗岩200 - 250 849 Ake Assi &菲(1975)
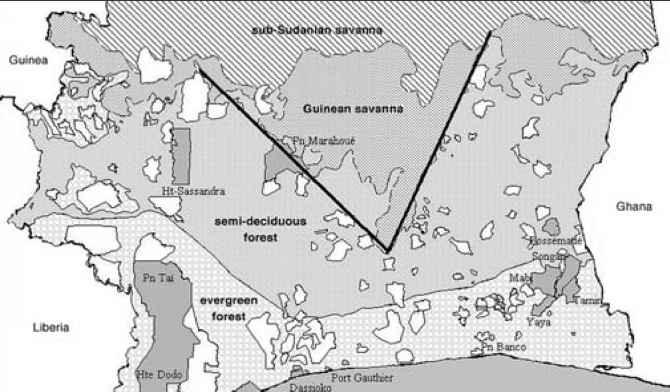
Moiofs»rtdantic Ocaan
图5.2科特迪瓦有四个地图主要植被类型(常绿森林,半落叶森林,几内亚热带草原,sub-Sudanian稀树大草原),森林保护区(白色多边形),13森林研究(深灰色多边形)。大胆的v型线表明稀树大草原入侵在森林地带,或V-Baoule。
不完整的物种列表。因此,在东南部森林面积(Mabi、Tamin Songan和亚亚),十六2公里,以前通过森林管理服务(SODEFOR-GTZ),用于物种库存。在高级渡渡鸟,由于缺少专门清除痕迹,我们随机分布的几个网站的库存,在预先存在的路径(Kouadio 2000)。这些库存是伴随着所有物种的库存示例图(Kouassi 2000)。我们都知道,物种列表不是详尽的分析了研究。结果是,我们的讨论都是基于知识的当前状态的植物象牙海岸的森林。
森林的物种列表使用S0renson成对比较的相似指数(1948)。这允许我们区分森林更相似的物种组成和那些更不同。指数的最大值(100%)表明两个森林相同的物种组成。最小值(0%)表明,两种森林没有共同之处。
随后13森林和2126类群的分类进行了利用对应分析(Jongman et al . 1987年),根据物种的物种列表的存在与否。任命分数与以下环境参数:经度和纬度,土壤和底土,一年一度的水分亏缺和总年降雨量。森林的群体相比,对应分析产生的成对使用S0renson相似性指数。的类群中发现所有的森林都被认为是最常见的物种象牙海岸的封闭的森林。清点所有的森林的分类单元相同的组,但没有发现任何其他森林群,被认为是典型的集团,他们被发现。
结果
13森林植物区系多样性的研究包含150个家庭,857属,2126种。的624个物种中发现只有一个森林,而41个物种被发现在所有的森林(表5.2)。物种的数量由森林共享是反向的
表5.2是常见的分类单元13森林,或类群特征的四个森林组织(图5.4)。
物种共有13森林
Aganope leucobotrya Agelaea paradoxa Agelaea pentagyna Aidia genipiflora Alstonia boonei Amphimas pterocarpoides Baphia nitida Baphia下毛竹Buchholzia coriacea Bussea occidentalis Calycobolus非洲
可乐nitida木香之后
Craterispermum caudatum Culcasia barombensis Diospyros soubreana Funtumia africana Glyphaea短Grifonia simplicifolia Klainedoxa gabonensis Landolphia物种Landolphia owariensis Microdesmis keayana Myrianthus arboreus Myrianthus libericus Napoleonaea vogelii Nephrolepis biserrata Neuropeltis acuminata Ochthocosmus非洲物种Palisota Parinari excelsa Piptadeniastrum africanum Psychotria peduncularis Pycnanthus angolensis毒毛旋花子gratus马针尾部Treculia africana Ventilago africana Xylopia quintasii Xylopia摘要
半落叶森林(我组)
Acroceras gabunense
Aneilema umbrosum
Bridelia atroviridis
Clerodendrum polycephalum
Cyrtococcum chaetophoron
山蚂蝗属adscendens var.robustum
Dichapetalum madagascariense var。
madagascariense
Diospyros abyssinica
尤金尼亚tabouensis
Grewia carpinifolia
非洲桃花心木grandifoliola
Lagenaria breviflora
Landolphia landolphioides
Melochia melissifolia
Mischogyne elliotianum var. glabra
Psychotria kitsonii
Psydrax manensis
Simirestis dewildemaniana
本文结合congolana
本文结合splendens
Telfairia occidentalis
牡荆ferruginea无性系种群。ferruginea
沿海森林(组2)
Ancistrocladus barteri Bulbophyllum imbricatum Eugenia whytei泽兰属植物microstemon Heterotis rotundifolia Salacia pallescens Salacia whytei Tapinanthus belvisii
西南森林(第三组)
Angraecum podochiloides Anthoclitandra nitida Bertiera fimbriata Bolbitis heudelotii Brieya下方Cercestis ivorensis Clappertonia小Dalbergia albiflora Delpydora股薄肌Didelotia brevipaniculata Drypetes klainei藤黄属植物granulata Gilbertiodendron robynsianum Gynura sarmentosa Lomariopsis俄罗斯Mapania小Millettia lucens Mussaenda landolphioides Pauridiantha hirtella Polystemonanthus dinklagei Premna grandifolia Psychotria subglabra Renealmia maculata Scleria vogelii卷柏杂色的马icaja Tarenna股薄肌Trichilia heudelotii牡荆ferruginea
东南部森林(第四组)
Aframomum alboviolaceum Buforrestia mannii Cecropia peltata木香englerianus Crotonogyne craterviflora Friesodielsia enghiana Guibourtia copallifera Guibourtia tessmannii Licania elaeosperma Marantochloa菲利普Memecylon polyanthemos Rutidea dupuisii无性系种群。occidentalis Sabicea变色与森林的数量相比(图5.3)。九个家庭都由50多个物种(表5.3)。茜草科、大戟科是最speciose家庭。最富有的属分别Psychotria和榕树与43岁和34个物种;他们榜首的六个属由至少20种(表5.3)。
银行之间的相似性指数最低(29%)和Marahoue(表5.4),它揭示了这两个国家公园的植物种类不同。之间的相似性指数最高(77%)Songan森林保护区和Tamin森林保护区,这最相似。最高的价值观也发现森林之间属于同一个地理区域(大/高级渡渡鸟,Songan / Tamin Mabi / Songan Tamin /亚亚)或森林之间的东南部和西南部的森林(高级渡渡鸟/ Mabi高级渡渡鸟/亚亚)。相似性指数变化从38% (Dassioko / Marahoue)到53% (Bossematie / Songan)与同一经度森林,但不同的纬度。
第一和第二轴的对应分析解释31%的物种组成的变化。44%的物种组成的变化是由六个环境变量来解释。四组的森林可以区分(图5.4)。组我,由半落叶森林(Bossematie,上流社会的Sassandra, Marahoue),特点是高水分亏缺(图5.5)和高纬度(图5.5 b)。沿海森林(银行、Dassioko、港口附近和Monogaga)第二集团,它的特点是一个纬度低,中间降雨,发生在第三纪沙土(图5.4)。第三组对应于西南部的森林(高级渡渡鸟,Tai),而第四组对应的森林东南(Mabi, Songan Tamin,亚亚)。第三和第四组
表5.3最常见的家庭和属13森林。每个显示家庭与50多个物种,属由至少20个类群。Psychotria主要草本和无花果属包括主要灌木。属Salacia Combretum,马和Dichapetalum lianescent。
表5.3最常见的家庭和属13森林。每个显示家庭与50多个物种,属由至少20个类群。Psychotria主要草本和无花果属包括主要灌木。属Salacia Combretum,马和Dichapetalum lianescent。
家庭 |
物种(#) |
属 |
物种(#) |
茜草科 |
260年 |
Psychotria |
43 |
大戟科 |
107年 |
热带榕属植物 |
34 |
蝶形花科 |
86年 |
Salacia |
28 |
夹竹桃科 |
77年 |
Combretum |
27 |
兰科 |
76年 |
本文结合 |
21 |
番荔枝科 |
70年 |
Dichapetalum |
20. |
苏木科 |
69年 |
||
桑科 |
52 |
||
翅子藤科 |
51 |
最大的植物区系多样性中心在科特迪瓦的低地。他们的特点是一个中间纬度位置,相当高的降雨和低水分亏缺(图5.4)。他们从根本上的不同类型的底土。西南的底土本质上是花岗岩,而东南由片岩组成。
一年一度的水赤字和纬度的位置是最好的环境因素与第一轴(图5.5、表5.5)。第二轴与三级沙土有很强的正相关,与强烈的负相关片岩(表5.5),并与纬度弱负相关(图5.4 b)。之间的相似性系数组森林产生的对应分析总是高于50%。这表明这些团体的森林有许多物种共同之处。西南和东南组织相似性指数最高(65%)。七十二种是森林的特征组:22个物种是典型的半落叶森林,8沿海森林,29日为西南森林和13东南(表5.2)。
表5.4矩阵相似性森林使用S0rensen指数(1948)。括号里的数字对应的累积的两个森林物种丰富度进行了比较。相似性指数大于50%(表明两种森林50%以上的共同点)是强调以粗体和大于60%。
表5.4矩阵相似性森林使用S0rensen指数(1948)。括号里的数字对应的累积的两个森林物种丰富度进行了比较。相似性指数大于50%(表明两种森林50%以上的共同点)是强调以粗体和大于60%。
森林 |
银行 |
Bosse-matie |
Dassioko |
上流社会的Sassandra |
附近 |
Mabi |
Mara-houe |
Mono-gaga |
Songan |
大 |
Tamin |
|
Bossematie |
(1382) |
|||||||||||
Dassioko |
(1495) |
(1328) |
||||||||||
上流社会的Sassandra |
42 (1618) |
(1447) |
48 (1561) |
|||||||||
高级渡渡鸟 |
(1685) |
(1518) |
(1632) |
(1751) |
||||||||
港口附近 |
(1482) |
(1313) |
(1427) |
49.5 (1546) |
(1617) |
|||||||
Mabi |
(1417) |
(1249) |
(1363) |
(1482) |
(1553) |
(1348) |
||||||
Marahoue |
(1250) |
50 (1082) |
(1195) |
(1318) |
(1385) |
40 (1182) |
(雷竞技csgo1117) |
|||||
Monogaga |
(1635) |
(1467) |
(1581) |
(1700) |
(1771) |
(1566) |
(1502) |
(1335) |
||||
Songan |
(1367) |
(1200) |
(1314) |
(1433) |
(1504) |
(1299) |
(1235) |
(1067) |
(1453) |
|||
大 |
(1625) |
(1457) |
(1571) |
(1690) |
(1761) |
(1556) |
(1492) |
(1325) |
(1710) |
(1443) |
||
Tamin |
(1289) |
46 (1121) |
(1235) |
(1354) |
(1425) |
49 (1220) |
(1156) |
(989) |
(1374) |
(1107) |
(1364) |
|
亚亚 |
(1392) |
(1224) |
(1338) |
(1457) |
(1528) |
(1323) |
(1259) |
(1092) |
(1477) |
(1210) |
(1467) |
(1131) |
变量 |
轴1 |
轴2 |
||
纬度 |
0.70 |
* * |
- 0.61 |
* |
经度 |
- 0.38 |
ns |
- 0.38 |
ns |
降雨 |
- 0.39 |
ns |
0.45 |
ns |
水分亏缺 |
0.70 |
* * |
0.15 |
ns |
花岗岩 |
0.14 |
ns |
- 0.04 |
ns |
沙子 |
- 0.11 |
ns |
0.90 |
* * |
片岩 |
- 0.02 |
ns |
- 0.80 |
* * |
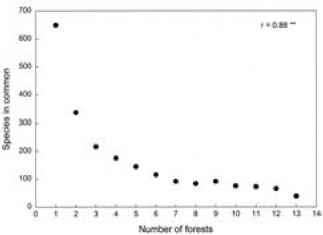
图5.3的物种数量之间的关系的共同点和森林的数量比较。斯皮尔曼等级相关系数。
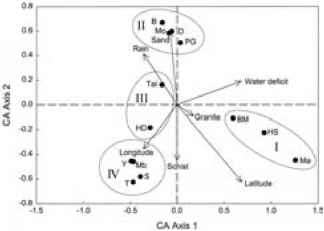
图5.4配合图13象牙海岸的森林,显示他们的立场在第一两个轴的对应分析。森林网站显示了符号,箭头和环境因素。森林被安排在四组。表5.1中给出缩写。
图5.3的物种数量之间的关系的共同点和森林的数量比较。斯皮尔曼等级相关系数。
图5.4配合图13象牙海岸的森林,显示他们的立场在第一两个轴的对应分析。森林网站显示了符号,箭头和环境因素。森林被安排在四组。表5.1中给出缩写。
讨论
植物区系多样性
9700平方公里的森林纳入本研究,代表了不到50%的森林覆盖的国家在1985年之前(戴维斯et al . 1994年)和不到3.6%的国家领土。然而,2126种维管植物被发现在这些森林,它代表总数的58%(3660种)象牙海岸的植物(戴维斯海伍德& 1994)。这表明13森林造成很大程度上的植物科特迪瓦。高贡献可以用这一事实来解释所有主要在科特迪瓦低地森林类型都包括在这项研究。剩下的42%的科特迪瓦植物在热带稀树草原,高地常绿的森林森林,森林岛屿,画廊,花岗质穹顶和人类环境。
这些森林的物种丰富度类似于某些国家如贝宁、利比里亚、塞内加尔和多哥。雷竞技手机版app它比塞拉利昂丰富的植物是评估在1700种(戴维斯海伍德& 1994)。它代表了近三分之二的多样性几内亚和加纳。植物的相对缺乏在贝宁、多哥和塞内加尔可以归因于他们的低降雨量和他们有限的森林覆盖。利比里亚是港口前冰川森林保护区(2000年莫理,Wieringa &要隘第6章)。利比里亚和塞拉利昂的相对较低的丰富性可以归因于这些国家缺乏知识的植物。雷竞技手机版app高级渡渡鸟是最富有的森林科特迪瓦至少有906种。
茜草科(260种)和大戟科物种(100种)是最富裕的家庭。在国家层面,这些森林分别为85和
这些家庭的75%的物种丰富度(Ake Assi 1984)。在几内亚热带稀树草原的代表蝶形花科,103种(1993年Kouame,禁止1995),在封闭的森林并不罕见,占86种。
Dioncophyllaceae的家庭,Hoplestigmata-ceae、Medusandraceae Octoknemataceae, Pandaceae和木果树科13中森林,并代表近70%的地方性家庭Guineo-Congolian地区1986(白色)。众多流行Guineo-Congolian属如Afrobrunnichia、Amphimas Anopyxis, Anthonotha, Antrocaryon, Aubrevillea, Buchholzia, Calpocalyx, Chidlowia, Coelocaryon,枚考拉,Crotonogyne, Cyclodiscus, Decorsella, Didelotia, Discoglypremna, Distemonanthus, Duboscia, Heckeldora, Hymenostegia, Gilbertiodendron, Grossera, Monocyclanthus, Ophiobotrys, Tieghemella和Turraeanthus也被发现在这些森林。
森林块之间的关系
在森林类型的区别是由水的可用性,这是总数的组合降雨量,雨量分布在今年,土壤的持水量。
物种组成最大的区别在我们之间的配合分析是一方面,半落叶森林,常绿和沿海森林另一方面(图5.4)。半落叶森林降雨明显变小了,全年分布是不均匀的。虽然沿海森林有一个类似的降雨和其他两个常绿林组高,植被明显降低水的可用性,因为高水分亏缺和沙质土壤持水量较低。两者之间的分离更微妙的常绿森林组织主要是由西南地区降雨量高引起的。
它ei
我'mi” |
VUi•助教 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
■一个 |
交通 |
V |
r |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
一个 |
Witv doUfri M ^
图5.5 A)之间的关系的第一轴分数森林和水赤字,B)的第二轴分数森林和纬度的位置。的回归,系数确定,显著性水平。 物种发生在只有一个13森林和只有2%的物种发生在所有网站。 森林植物区系相似性的两个不同29至77%,平均在50%左右。一般来说,植物区系相似性与网站之间的距离,环境条件的相似性,或两者的结合。所有三例在这项研究中被发现。例如,邻近森林Bossematie而且Songan比例近50%的物种,尽管在环境中明显的区别。东南和西南地区的森林,相距400公里有一个植物区系相似性高,因为高相似度的水可用性。细微的差别在降雨和水分亏缺这两个区域被底土的自然衰减。东南的粘土土壤(Perraud & Souchere 1970)砂质土壤的保水能力大于西南(De Rham 1971),从而为这个地区的低降雨量补偿。因此,植物经历类似的水可用性在这两个区域。最后,森林地理上接近和相似的土壤和气候条件,对Mabi一样,Songan, Tamin,亚亚,植物区系相似性最高(表5.4)。 古气候历史两个森林可能是另一个原因是相似的。假设,在极端的西南部和东南部森林科特迪瓦的两位前冰川森林保护区(Guillaumet 1967年,Wieringa &要隘第6章)。潮湿的森林物种可能在这些生物避难所干冰川时期幸存下来。许多物种的地区或大陆分离的分布模式人口在这两个两个森林块(霍蒙格林等人第7章)。总结,象牙海岸的森林之间的差异都是由一个复杂的相互作用raybet雷竞技最新气候、土壤和历史。 尽管银行有很高的降雨,相对远离海岸(13公里)还一直与沿海森林分组(图5.4)。高降水量由砂质土壤的持水量低,导致低水的可用性。决定这个植物区系组最重要的环境因素是土壤;第三金沙下森林,导致较低的水和养分的可用性。 每个森林组具有良好定义的物种群特征(表5.2),例如,非洲桃花心木grandifoliola在半落叶森林,Ancistrocladus barteri在沿海森林,Mapania西南部小森林,和两种Guibourtia东南部森林。令人吃惊的是,几乎三分之一的 确认我们想感谢SODEFOR允许我们在森林保护区开展研究。我们感谢克劳德·阿玛尼,Jean Assi Saturnin Dougoune,火绒Fofana和帕特里斯·Mabea协助数据收集。我们感谢教授Laurent Ake Assi识别分类单元,和特拉奥雷Dossahoua Frans Bongers评论,这让我们改进这一章。 J.J. Wieringa和l .要隘  生物多样性热点地区在西非;模式和原因 介绍西非的雨林被指定为世界上的生物多样性热点地区之一(迈尔斯et al . 2000年)。这些森林从多哥扩展到塞内加尔,被称为上几内亚森林1983(白色)。上几内亚森林分开其余的非洲热带雨林女兵的差距;在多哥和贝宁林地草原延伸从北到几内亚湾。 上几内亚森林港口大量流行的植物和动物物种。据估计,约2800维管植物物种可以在上面找到几内亚森林,其中22%是该地区特有的(Jongkind,第11章)。这些森林正在迅速消失,因为伐木活动,培养,和转换成种植园(城主等人第二章)。对于一个有效的保护政策,信息需要稀有和特有物种的分布上几内亚、以及他们集中的地方。许多的问题热带国家雷竞技手机版app是这种植物背景信息很少,或高度分散。快速生成必要的信息可以进行植物调查,筛选选定地区的物种组成。霍桑和Abu-Juam(1995)使用这种方法在加纳。基于样本块,系统地分布在整个森林地带,他们能够划分区域高份额的特有种。然而,这种方法是劳动密集型,仅仅在限制植物区系和地理范围覆盖着一支训练有素的植物学家。另一个选择是使用现有的标本收藏作为数据源。植物收藏有一个好处,那就是它们提供一个现有的信息来源,涵盖了大地理范围,他们可能会被正确识别。后者是重要,特别是稀有物种不易被树观察员。缺点是,收集努力不是均匀分布在该地区。 几个作者使用了小群体的分布模式植物或动物来表示领域具有高生物多样性(汉密尔顿Aubreville 1949、1962、1976、Grubb 1982, Sosef 1994年,洛维特et al . 2000年)。这些研究通常点在上几内亚的三个方面:内部
利比里亚的多样性中心周围山宁巴,帕尔马斯角之间的边境利比里亚、科特迪瓦和加纳西南部三分角及其周边地区。热点的确切位置可能不同,不同分类群的关注(保护国际2001)。一些生物多样性分析更多的物种存在(例如城堡1993,林德2001),但他们有一个相当低分辨率。到目前为止一个详细的定量分析,基于大量的物种因此缺乏对于上层几内亚。 环境和历史因素影响物种丰富度的空间变化。物种丰富度是沿着环境梯度的变化而变化的降雨量(大厅&斯1976,库里1991,1993 O ' brien),海拔(1973年大厅),土壤肥力(霍尔&斯1976,休斯顿1980)。一般来说,更好的现场条件导致初级生产力增加,更多的个人和利基市场,因此,物种丰富度更高。物种丰富度可能会减少在富有成效的网站,导致一个单峰丰富性和网站生产力之间的关系(1973年污垢,休斯顿1979)。 当前植物区系模式是强烈的大规模气候扰动在过去。整个第四纪气候波动而起伏的热带雨林面积(1991年汉密尔顿&泰勒)。在干燥和凉爽的冰川时期,降水量减少,热带雨林简约小补丁。随着降雨水平很低在非洲与其他大陆相比,冰河时期的影响一直觉得这里比其他地方更强烈(理查兹1973年,莫理2000)和它的印记很可能持续了更长时间。证词稀树大草原植被的动态变化残遗热带雨林区内在科特迪瓦(Gautier & Spichiger 3章),并观察Guineo-Congolean森林树木的化石在埃塞俄比亚(Bonnefille & Letouzey 1976)。一些物种不能跟上快速扩张,并仍在一个狭窄的范围约前森林保护区。陆地森林草秋海棠mildbraedii,例如,有一个分离的分布,有两个孤立的种群在科特迪瓦和加纳,是除了其主要1800公里,更加广泛,分布在非洲中部(霍蒙格林等人第9章,Sosef 1994)。 本章着重于模式和植物生物多样性热点地区在西非的原因。热点被定义为高的中心丰富的稀有和特有的物种。这一章探讨了如何以及在多大程度上标本收藏可以用来定义生物多样性的热点地区。然后有关生物多样性环境和距离假定前森林保护区,重环境的重要性和历史在当前生物多样性的分布。 方法 数据收集第二版的基础上西非洲热带的植物(1958 b Keay 1954年,1963年,1968年玫瑰,1972),库存在加纳(大厅&斯1981,霍桑1995 a),分类修订、和新标本收藏,我们做了一个编译超过1000种,珍贵、特有的封闭森林上几内亚(见第11章Jongkind & Wieringa)。所有生命形式包括(树木、灌木、藤本植物、草本植物、寄生虫、腐生菌和附生植物)。640个物种被选为一个名单来分析生物多样性模式。谨慎,包括物种来自不同的家庭,并包括与不同物种分布模式或生态。这些640个物种,植物标本的标本被进入c。510种,这是辅以分布数据分类修正c。130个物种。 我们进入到一个数据库中所有的植物标本提供收集从塞内加尔到多哥。对于一些非稀有物种,我们也包括植物标本的标本收集到较低的几内亚。我们进入所有标本的标本Vadense(荷兰瓦赫宁根),
比利时国家植物园(Meise、比利时)和基尤植物园(丘,英国)。此外,我们添加了集合从科特迪瓦出现在日内瓦标本数据库(艺术学校等查顿Botaniques de la城镇de日内瓦,瑞士)(只有到数据属于日内瓦)、科特迪瓦和几内亚的集合出现在巴黎标本数据库(国家d国立博物馆,巴黎,法国),和100个物种的集合出现在大学的植物标本加纳(Legon、加纳)。对许多非物种标本几内亚也进入低。总共从西非数据库包含48000条记录,其中超过12500条记录对应于珍稀特有植物从我们的名单。 数据库涵盖了所有主要的植物标本室收藏从西非,除了巴黎标本是重建我们的数据输入期间关闭。对于我们的地区,我们希望巴黎标本包含主要收集来自科特迪瓦和几内亚。因为我们有丰富的藏品从科特迪瓦从其他来源,真正的问题是缺乏来自几内亚的数据。引用集合由骑士在几内亚追溯使用骑士(1911、1920)。几内亚虽然我们的结果会更准确的数据,很可能他们是足够好的那些地区森林物种的最大数量是预计(Fouta Djalon,太Ziama太宁巴)。识别不同植物标本室的标本被c . Jongkind反复核对,有充足的知识分类的西非森林物种。术语更新使用霍桑& Jongkind (2004)。 6.1箱。Species-collection曲线允许区域之间的比较,可以构造一个species-collection曲线,以同样的方式作为一个物种-区域曲线。为每个网格单元的半度半度经度纬度,我们随机集合,和绘制累计数量的物种,累积的集合(图6.3)。这样100 species-collection曲线构造和随后的平均。给定区域中发现的物种数量的增加与收藏的数量,直到达到渐近线(Colwell &科丁顿1995,1998年的怪兽Steege)。所以我们安装每个细胞渐近曲线通过的数据,根据方程Sn = (Smax * n) / (c + n), Sn是物种的数量在一个示例(n)集合,Smax估计最大的物种数量在一个给定的区域,和c是一个常数(图6.3)。对于这种分析我们在集合属于我们640个物种的名单。 生物多样性的估计更加准确,许多标本收集了(N)相对于物种的数量(抽泣)。我们认为我们的生物多样性估计是相当满意的,如果一个细胞的比率N /呜咽,大于1.5。如果N /呜咽,< 1.1,那么生物多样性的估计相差很大,和太不可靠的使用。我们只通过数据拟合曲线,如果我们有八个或更多集合。八细胞尽可能多的物种被发现集合,所以曲线拟合是不可能的。另一个七细胞N /抽泣是小于1.1。这与回归结果细胞的数量减少到154。  4 + 1-1-1-1 - n nuG 4 xo ud nr ioooc 4 + 1-1-1-1 - n nuG 4 xo ud nr ioooc GotoctxroiH图6.3 Species-collection曲线细胞,包括银行森林,科特迪瓦。species-collection曲线是由随机图集合从总收集池在55 * 55公里范围之内。累计物种数量绘制的累积数量集合(圈)。随后回归曲线拟合是通过数据使用一个渐近曲线。通过使用曲线,可以计算预测物种丰富度(虚线),和物种的最大数量(Smax)可以被估计。 如何分析生物多样性模式? 描述大规模模式生物多样性,上层几内亚应该分成同等大小的区域。应该选择什么网格大小来描述这些模式最好?一个实际的考虑,应该有足够的集合中每个单元格进行有意义的比较。更理论的考虑是,网格的大小不应太小,其他本地站点条件可能会否决大规模的照片。作为一种妥协,我们选择了一个gridsize 0.5 X 0.5度。 植物有几个优势集合;他们提供现有的信息来源,可能是正确的确认(如果标本不是太小,如果物种组最近修订的),并提供永久记录,总是可以重新核对。缺点是样本收集不分层或随机的方式进行。因此,抽样努力或随机分布是不均匀的,和测量物种密度高的区域往往配合地区收集的高强度(Nelson et al . 1990年)。同样的标本来自西非的数据。如果西非分为相同细胞的0.5 x 0.5度,还有一个大集合的变化强度;为我们的物种名单收集每个细胞的数量从0到1303不等。观察到的每个细胞的物种数量增加曲线的方式的集合(图6.2,第二学位多项式,r2 = 0.96, P < 0.001, n = 329)。网站似乎有大量藏品的物种,但这很大程度上可以归因于更高的采样工作。 曲线的形状就像物种-区域曲线(加斯顿1996年),和底层采样机制有点类似;区域或集合抽样数量越大,物种被发现。允许区域之间的比较,可以构造一个species-collection曲线,以类似的方式一个物种-区域曲线是由(框6.1)。species-collection曲线几种生物多样性的措施。S 50表明物种的数量在50集合是由640年的名单珍稀特有植物(框6.2)。Smax估计最大数量的物种从我们的名单,可以发现在一个单元中(框6.2)。最后,rarity-weighted物种丰富度(Srw)表明地区大量的稀有和特有物种(框6.3)。这三个生物多样性措施地图已经在ArcView使用反距离加权插值法。细胞的曲线拟合是不可能的(N =抽泣),或使用,被认为是不可靠的(N /抽泣< 1.1,参见6.1)并不包括在 6.2箱。S50 Smax;两个物种丰富度的措施 不同网格细胞的多样性可以相比,使用species-collection曲线。图6.4显示了这样一个species-collection曲线三个不同的网站。Tabou,物种数量增加速度比大的收集数据,进而远比银行更为多样化。因此Tabou最大的多样性。然而,多样性是逆转的排名,当一个人只是比较物种的总数(银行192种,大的160年,Tabou 119)。原因是在银行集合已经远远超过其他两个网站。根据我们计算在银行84%的实际发生的物种也被发现,而对于Tabou这是仅为38%。因为它会太复杂,许多species-collection曲线进行比较,我们使用作为生物多样性措施物种的总数在一个标准的集合。帮助下,为此我们计算回归方程,预期的物种数量在50岁(S50)集合。的参考价值50接近每个网格单元的集合数中值(值= 35范围8 - 1303),因此不需要做太多的推断。 S50 can vary from 1 (if all 50 collections in a cell belong to the same species) to 50 (if all 50 collections belong to 50 different species). 如果我们将无限的集合,我们将方法物种的最大数量(Smax)细胞。Smax表明有多少稀有物种出现在一个区域。Smax和S50强有力的指数关系(r2 = 0.96, P < 0.001);因此更大的S50, Smax大。S50的高层小S50的增加将导致
im isa im isa CMnll 图6.4安装物种集合曲线50 X 50公里周边地区Tabou(实线),大(小折线)和银行(虚线)。为每个站点的实际观察到的物种数量之间的面积表示括号。 不均衡地在Smax大幅增加。Smax因此是一个生物多样性的措施,提供了一个大分辨率高水平的物种丰富度。Smax的一个缺点是需要做出推断。在偶然的情况下,这可能会导致一个实数的高估或低估物种出现在该地区。 插值分析。插入生物多样性值只显示森林地带。 生物多样性和环境之间的关系水可用性、高度和土壤肥力的形状在很大程度上和植物群落的组成结构。环境变量用于分析降雨(毫米/年)、土壤持水量(通车,在毫米水土壤/ m),高度(米),土壤肥力(cmol中钙、镁、钾、土壤阳离子每公斤),土壤pH值和阳离子交换量(CEC %)。降雨地图创建基于该地区578个气象站的编译。数据在土壤肥力和土壤持水量的计算是基于联合国粮农组织非洲地图,和定量评估的土壤化学分析资料(Batjes 1997)。为每个网格单元平均土壤肥力进行了计算,基于相对覆盖不同的土壤类型,和他们的平均土壤肥力。同样,对于每一个网格单元平均土壤持水量计算,基于相对覆盖的土壤类型,深度和纹理。假定,砂质土壤的持水量75毫米/米,100毫米/ m肥沃的土壤,粘质土壤的125毫米/米。环境的元数据的详细分析,见第9章。所有空间分析进行了使用ArcView (Esri Inc .)。 S50和Srw相关环境变量用斯皮尔曼等级相关和逐步多元回归。除了这些非生物因素,也距离冰川森林避难所被用作提出的三个因素进行了分析。在某种程度上,似乎循环与特有物种的丰富性的距离提出森林保护区,特有现象和庇护理论紧密相连。然而,这些森林避难所的存在和位置一直被假设的基础上主要花粉核和古气候数据(Maley 2001),和与分离的物种分布之间上下几内亚(Guillaumet Aubreville 1949年,1967)。在较小程度上已经发生的基础上,物种分布范围受到限制。 6.3箱。Rarity-weighted物种丰富度S50和Smax是物种丰富度的一般措施,不考虑物种的“特殊”。出于保护的目的,人们可能会有兴趣知道罕见的物种集中,或有一个有限的分布。为此我们计算rarity-weighted物种丰富度(Srw)。这是物种多样性的措施,使体重和许多珍稀特有植物细胞。每个物种的数量进行加权half-degree网状细胞的发生。常见的物种,在100年发生half-degree网状细胞接收因此1/100的加权分数每个细胞的发生。稀有物种的一半发生在4度网状细胞接收权重评分为0.25。rarity-weighted物种丰富性然后权重之和的所有物种发生在一个细胞。然而,所有的物种都发生在一个细胞(Smax),只有一部分(SobJ采样。为了纠正这一点,我们增加加权测量Smax /呜咽例如,如果一个网格单元有10集合属于4种物种发生在1细胞,2在2细胞和1 4细胞,然后加权丰富的细胞= (1 X 1 + 2 X 0.5 + 1 X 0.25) = 2.25。 The predicted species richness (Smax) of the cell is 16, four times as much as actually observed. When correcting for sampling intensity, the rarity-weighted species richness equals rarity-weighted物种丰富性背后的基本原理是,在大规模(上几内亚),每一个物种都是同样重要的,接收的总重量为1。我们在分析包括640种,所以之和rarity-weighted丰富的物种在全球所有细胞= 640年。也因为一些物种发生上几内亚外,细胞在上层几内亚的总和是498。因为我们后来增加每个单元与Smax /抽泣,点的总量分配给上层几内亚细胞变成了1406。因为上几内亚有保护价值的498种点,我们新rarity-weighted物种丰富度的每一个细胞通过乘以0.354(= 498/1406),再到在原来的498分。 生物多样性的表6.1给出一个示例计算两种不同的细胞。在细胞只有几个,但是非常罕见的物种被发现。在B细胞更多的物种被发现,但他们也越来越普遍。虽然物种中发现了一个更少,其rarity-weighted丰富性是B的近四倍。 rarity-weighted丰富的细胞可能会有所不同从接近0(如果只有一个物种发现发生在许多网状细胞)到640(如果所有640种碰巧在同一细胞和其他地方)。这种多样性测度的一个有趣的特性是,它把植物区系的细胞区域,上层Guinea-wide视角。如果一个细胞rarity-weighted丰富的它可能表明它只包含10个物种被发现,和其他地方。另外,它可能包含20个物种,只出现在这里,在另一个细胞在世界的每个角落。当然每个细胞都有一个不同的保护值不同的物种,但是rarity-weighted丰富是一个很好的测量细胞的总价值的个别物种的保护和长期生存。 表6.1的例子的计算rarity-weighted物种丰富度与几个虚构的细胞但非常罕见的物种(细胞),并与许多细胞常见的八种物种(细胞B)。它是表示有多少个人在a和B细胞,发现在他们发生,多少细胞及其权重分数(重量)。 表6.1的例子的计算rarity-weighted物种丰富度与几个虚构的细胞但非常罕见的物种(细胞),并与许多细胞常见的八种物种(细胞B)。它是表示有多少个人在a和B细胞,发现在他们发生,多少细胞及其权重分数(重量)。
 图6.5分布的植物标本收藏(点)of640稀有和特有的森林物种在西非。潜在的森林区域阴影。  图6.6抽样强度(100 X哭泣/ Smax) of640稀有和特有的森林物种在西非。 图6.5分布的植物标本收藏(点)of640稀有和特有的森林物种在西非。潜在的森林区域阴影。 图6.6抽样强度(100 X哭泣/ Smax) of640稀有和特有的森林物种在西非。 结果 收集工作;地图上的白色斑点在哪里? 我们大部分的集合名单的森林物种局限于非洲西部和南部确实,密切关注森林区域(图6.5)。首都附近的收集工作一直特别高(弗里敦蒙罗维亚,阿比让)和植物研究站附近(见附录5西非的地图)。到目前为止最多的集合了阿比让以西。银行国家公园和研究站在Adiopodoume属于half-degree广场,导致1303年集合属于190个物种从我们的名单。细胞阿比让以北,包括Yapo森林和森林泰克,包含生态站在Ta的细胞“分数很高。在利比里亚,旁边蒙罗维亚太宁巴周围地区的环境已经被收集。在塞拉利昂三细胞相对许多集合:朝鲜半岛(包括首都弗里敦),在Njala林业研究站,Kambui山保护区。在加纳的Ankasa森林保护区和Atewa范围是最高的细胞数量的集合归因于我们的物种名单。 如果我们观察到的物种数量除以预计最大的物种数量,我们得到一个衡量抽样强度。抽样强度非常低(< 20%)利比里亚东南部(图6.6)。这个区域包含一个最大的现存森林块在西非,也收到大量的降雨。可能因此港口一个富裕和未知的植物,这绝对值得进一步关注。塞拉利昂,东南部和西南部加纳(Ankasa以北和角三分Bia国家公园)是另外两个潜在的物种丰富度高的大面积但很少物种采样在我们的数据库中。规模较小等领域引人注目的是,东南大国家公园的一部分,国新办 森林保护区是如此糟糕了,我们甚至不能估计抽样强度。 热点的多样性 有一个南北梯度物种丰富度,伴随着降水梯度;S50(图6.7)和SmD(图6.7 b)增加来自西非的居民涌向海岸。带高稀有和特有物种丰富度约50 - 100公里的内陆,从塞拉利昂,利比里亚贯穿西南科特迪瓦,然后向Sassandra消失。这个范围的高潮在于利比里亚和西南科特迪瓦。额外的发达地区,东部的皮带被发现在阿比让和Ankasa(加纳)。第二带高稀有和特有物种丰富度可以找到更多的内陆山区面积太宁巴在几内亚和扩展Mt Ziama山区周围地区在科特迪瓦人。在加纳Atewa范围可以被视为一个走在时代前端的飞地丰富的山区地带。 一些地区种类匮乏地区相对丰富的直接环境或其他领域相比,他们相同的纬度。例子是沿海地区在几内亚比绍、塞内加尔和Casamance地区的Fouta Djalon在几内亚、塞拉利昂的弗里敦半岛,上流社会的Sassandra森林和Comoe国家公园在科特迪瓦(图6.7)。 模式在rarity-weighted丰富性相媲美的物种丰富度(图6.7摄氏度),尽管丰富的山地地区与沿海地区相比变得更加明显。例子太宁巴,太Ziama,太Tonkui Atewa范围。在沿海丰富的乐队也最潮湿的森林变得更加明显,这是对Tabou和格林维尔表示非常高的价值。 生物多样性与环境 什么环境因素引起物种丰富度高吗?S50显示与降雨关系曲线; 一个S50  一个S50
 图6.7生物多样性西非的地图显示)的物种数量在50岁(S; o)集合,B)的最大估计稀有和特有物种的数量(Smax)和C) rarity-weighted物种丰富度(Srw)。生物多样性价值的细胞内插在整个潜在的上层几内亚森林地带。 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

. Two cells with a collection intensity larger than 500per cell are not shown.")

继续阅读:如何解释西非稀有和特有植物的分布
这篇文章有用吗?