The members of the hot club
The ability of birds and mammals to regulate their internal temperature allows them to survive in some hot environments. Other organisms can do so by behavioural or structural mechanisms which allow them to lose, or avoid gaining, heat. Some organisms, however, lack these mechanisms and their lifestyle or habitat means that they have had to join the 'hot club' and survive high internal temperatures.
What's hot and what's not?
The great majority of organisms live in environments where temperatures are in the range of 0-48 °C. There are few habitats where temperatures rise above 48 °C and fewer still where temperatures are constantly above 48°C. Temperatures below 0°C are more common, exposing organisms to freezing or the threat of freezing. How organisms cope with temperatures below 0 °C will be described in the next chapter. Few animals or plants can survive sustained exposure to temperatures above 48 °C for more than a short period. Such an ability might thus be a useful criterion for membership of the hot club. Birds maintain an internal temperature of 41 °C, only seven degrees below the cut-off point. Since they happily survive this temperature inside their bodies, the destructive effects must kick in rather soon as temperatures rise. The lethal temperature is only a few degrees above that which is optimum for their metabolism. Some proteins start to be denatured at 45°C and most animals die after exposure to 50°C for more than brief periods.
In hotsubtropical deserts, air temperatures regularly reach 50 °C and ground temperatures 70 °C, absorbing the baking heat of the sun. As we saw in Chapter 2, however, desert plants and animals either avoid the heat or have mechanisms which prevent them from heating up to lethal levels. Desert microorganisms which cannot avoid the heat or cool themselves down may have to survive exposure to these high temperatures. Along with the heat, however, is likely to come desiccation.Anhydrobiotic生物体在干燥状态可以阻力t much higher temperatures than when they are hydrated (see Chapter 3). Protein denaturation by heat involves the molecule unravelling as the increased kinetic energy breaks the weak bonds which maintain its shape. This cannot happen in the absence of water, andanhydrobiosisalso confers heat stability. Desert microorganisms are thus likely to survive high temperatures in a state of anhydrobiosis. Some anhydro-biotic animals (nematodes, rotifers, tardigrades and some arthropod larvae) and plants (resurrection plants and seeds) are also able to survive high desert temperatures in a desiccated state. These organisms use resistance adaptation - lying dormant until water, and lower temperatures, return.
Terrestrial hot springs and deep-seahydrothermal ventsare aquatic habitats and the organisms which inhabit them cannot rely on anhy-drobiosis to survive (since they cannot desiccate). The conditions are also fairly constantly hot and the organisms use capacity adaptation to adapt to, and thrive in, the hot environment. They are thus extremo-philes (thermophiles). There are no plants or animals that can survive these conditions which are constantly above 50 °C (or maybe just one animal, the Pompeii worm - see later). Membership of this hot club is restricted to microorganisms.
Hot club plants and animals
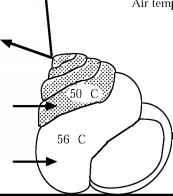
Desert animals which cannot avoid the heat and which do not have mechanisms to cool themselves down may reach temperatures (above 48°C) that qualify them for membership of the hot club. The small desert snail Sphicterochila boisseri inhabits the Negev Desert of Israel. In dry conditions, it becomes dormant on the surface of sand and rocks where it is exposed to the full heat of the sun. The surroundings of the snail reach temperatures as high as 65 °C on the ground and 43 °C in the air. The shell is very shiny and reflects much of the solar radiation and the snail retreats to the upper whorls of the shell, leaving an air pocket
95% of solar radiation reflected by white shell
Snail tissue
Air space
Air temperature = 43 C
95% of solar radiation reflected by white shell
Snail tissue
Air space
65 C
60 C
65 C
figure 4.2 Temperature relations of the desert snail Sphicterochila boisseri. Redrawn from Schmidt-Nielsen et al., 1971 and reproduced with the permission of the Company of Biologists Ltd.
which insulates it against heat gain from the ground. The snail's tissues, nevertheless, reach temperatures as high as 50°C, but it can survive this since its lethal temperature is between 50 °C and 55 °C (Figure 4.2). The animal is, however, dormant under these conditions and it is using a resistance adaptation. The snail does not desiccate appreciably and can maintain a water content of 76 per cent for several weeks under hot dry conditions.A desertant, Ocymyrmex barbiger, forages during the day when temperatures on the surface of the sand are as high as 67 °C. The ant can avoid the extreme heat by raising its thin body above the surface on its legs and by seeking out shade and cooler areas. In spite of this, it survives body temperatures in excess of 52 °C for short periods, although this is close to its thermal death point. Foraging during the day is risky for the ant and many of them die from heat stress, but it allows them to be first at the scene to scavenge on the carcasses of other insects that have died from the heat. They also avoid the attentions of lizards and other predators which cannot sustain activity during the hottest part of the day.
Desert plants cannot move away and avoid exposure to the sun and heat. Tidestromia oblongifolia is a herbaceous perennial which grows during the summer in Death Valley, California, at temperatures in excess of 50°C. It cannot grow in more moderate climates near the coast and is thus clearly heat adapted. Desert plants reduce heat gain by having a light colour and spines, hairs and waxy cuticles which reflect the sun's rays or shade the surface of the plant. Some plants (including Tidestromia) can cool down by having rapid rates of transpiration (which produces evaporative cooling at their surface). Desertsucculents(includingcacti), however, conserve their water and could not survive the water loss involved in evaporative cooling. Their internal tissues can reach temperatures above 60 °C and the plants can survive these temperatures for prolonged periods. One of the highest temperatures recorded in a live plant is 65 °C in a species of Opuntia (acactus).
很少有动物或植物生活在terrestrial geother-mal springs. Brine flies will land and feed on the algae growing in the springs but only where temperatures have cooled to below 40 °C. The flies lay eggs that hatch and the larvae which emerge also feed on the algae. The larvae die, however, if the flow of water in the spring changes so that they are swept away or exposed to water which is hotter than that which they can tolerate. A nematode (Aphelenchoides parientus) has been reported from a hot spring living at temperatures of 45-51 °C and insect larvae (chironomids) from hot springs at 49-51 °C.
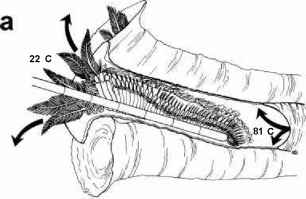
The water issuing from deep-sea hydrothermal vents can exit, superheated because of the highpressure at depth, at temperatures over 300°C. It rapidly cools, however, since the surrounding seawater is at about 2 °C. The animals associated with the vents live mainly in waters at around 30 °C. Perhaps the best animal candidate for membership of the hot club is the Pompeii worm (Alvinellapompejana, a small polychaete annelid) which lives on the walls of black smokers, the chimney-like structures which form by the deposition of minerals around some deep-sea hydrothermal vents. The worm builds a tube, in which it lives, on the walls of the chimney. Hot water from the vent flushes through the tube, cooling as it mixes with the cold water at its mouth. Probes inserted into the tube using a 'deep submergence vehicle', by Craig Cary from the University of Delaware and his coworkers, have measured temperatures averaging 81°C at the base of the tube and 22 °C at its mouth. When probes were positioned at the tail end of the worm, temperatures averaging 68 °C, and occasionally exceeding 81 °C, were measured. If these measurements represent the temperature of the worm itself, the Pompeii worm would not only be the most thermotolerant multicellular (and eukaryotic) organism known, but also one which can survive an unprecedented thermal gradient of up to 60 °C (Figure 4.3). Given the marked temperature gradient within the tube, however, the precise relationship between the position of the worm and that of the temperature probe is critical. The measurements were made using probes positioned by an operator in a deep submergence vehicle. The precise placing of the probe and the monitoring of its position in relation to the worm was therefore difficult. Craig Cary tells me that their most recent temperature measurements, however, confirm their earlier findings and that they have recorded temperature spikes up to 89 °C.
Another approach to determining the tolerance of the Pompeii worm is to examine the thermal stability of its enzymes and the other molecules and components which are vital for its survival. The worm's haemoglobin becomes unstable at temperatures above 50°C and the respiration of isolated mitochondria ceases above 49 °C. Denaturation of its cuticle collagen (the protein which is the main structural component of the body wall) occurs around 45 °C. These measurements, and the temperature optima of its enzymes, suggest that the worm is limited to temperatures below 50 °C, with an optimum body temperature of 30-35°C. Craig Cary has, however, isolated an enzyme from bacteria which are associated with the surface of the worm that has a temperature optimum of about 82 °C. It is difficult to relate these measurements on isolated components to the actual thermal
M-Vent Temperature Profiles
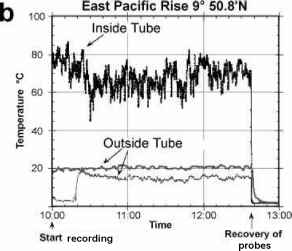
figure 4.3 Temperature recordings from the tubes of the Pompeii worm, Alvinella pompejana. The worm is about 6 centimetres long.
(a) Drawing showing the worm in its tube with a temperature probe in position. The temperatures shown are the maximum recorded during one deployment of the probe.
(b) Temperature recordings from inside the tube and two recordings from outside the tube, taken over a three-hour period.
Reprinted by permission from Nature (Cary et al., 1998) copyright (1998) Macmillan Magazines Ltd. Graphic kindly supplied by Craig Cary of the University of Delaware.
environment of the worm. In the intact animal, protective thermal mechanisms may operate (such as via molecular chaperones). If there is indeed a marked temperature gradient along the body of the worm, thermally sensitive functions may be centred in the cooler parts of the animal. There is clearly more to learn about this fascinating animal.
Hot club microbes
New Zealand, my adopted home, lies on the boundary of two of the greatcontinental plateswhich make up the surface of the Earth. Here,the Pacific Plateis drawn under the edge of theIndo-Australian Plate这一过程称为俯冲,形成能力lt line which runs for more than 600 kilometres through both the South and North Islands. The forces generated by thesubduction processhave created huge mountains (the Southern Alps) and regular earthquakes. As the Pacific Plate is dragged beneath the Indo-Australian Plate, it is forced deeper into the Earth, encountering hot temperatures which melt its rocks and convert them into molten magma. In places, this finds its way to the surface via cracks in the crust to form volcanoes. The active volcanoes of New Zealand, White Island and Mounts Ruapehu and Ngauruhoe, are part of the Ring of Fire - achain of volcanoesaround the edge of the Pacific. When water comes into contact with the hot rocks, it may emerge at the surface in the form of bubbling mud pools, geysers and hot springs at temperatures up to 101 °C. The area around the lake-side town of Rotorua in the central North Island is New Zealand's best known thermal area. The whole town is pervaded with the bad eggs smell of hydrogen sulphide gas (known affectionately by the locals as 'Soir de Rotorua') andone areawas named Whangapipiro ('an evil-smelling place') by the indigenous Maori people. Bubbling mud pools are one of the more unusual hazards to be encountered on the local golf courses. Hot springs are found throughout the world, associated withsubduction zonesand volcanic activity, with their greatest concentrations in the western USA (including Yellowstone National Park), Iceland, New Zealand, Japan, the Mediterranean countries, Indonesia, Central America and Central Africa.
There are no animals or plants in these hot springs, in which the water is too hot to immerse your hand, but these scaldingly hot waters are not devoid of life. The presence of bacteria is revealed by, often brightly coloured, films and stringy filaments. If glass slides are immersed in the water, they rapidly become colonised by bacteria. In the 1960s, Tom Brock, now retired from the University of Wisconsin-Madison, started isolating and culturing bacteria from the hot springs of Yellowstone National Park. He found many different types of thermophilic bacteria, including several new species which lived at temperatures in excess of 73°C. Large yellow masses of filaments, from which a bacterium identified as Thermus aquations can be isolated, grow in the hot springs of Yellowstone at temperatures of 75-80 °C. Some of the hot springs of New Zealand reach higher temperatures than those of Yellowstone because of their lower altitude. On a visit to New Zealand, Tom Brock was able to isolate bacteria at temperatures of around 100 °C. He could not, however, isolate any organisms from fumaroles (crevices in the walls of a volcano through which steam issues) in Italy and Iceland which produced superheated steam at temperatures much greater than 100 °C. It appears that the bacteria need the presence of liquid water and that the boiling point of water is the limit for life in terrestrial hot springs.
The presence of bacteria in boiling water is limited to hot springs where the water is at an alkaline or neutral pH. Acidic conditions are more difficult to cope with and the upper temperature limit for organisms in acidic hot springs is about 90°C. The high pressure in the deep sea allows water to emerge superheated from hydrothermal vents at temperatures above 300 °C. Bacteria growing near the vents have been isolated at temperatures of about 115 °C. These have been called hyper-thermophiles, extreme thermophiles or ultrathermophiles - all ways of saying 'boy are they hot!'. Some of these, such as Pyrodiotium and Pyrolobus, have optimum growth temperatures above 100°C (105°C). Hyperthermophilic bacteria have also been isolated from the production fluids of oil fields at temperatures up to 110 °C. Even hotter conditions, which nevertheless support bacterial growth, may be found in the deep subsurface. It is thought that the temperature limit for bacterial life may be up to 150 °C.
只有细菌can survive extreme thermophilic conditions. The upper limit for the growth of thermophilic fungi is around 60°C, the maximum growth temperature of the alga Cyanidium caldarium is 57 °C and protozoa have been grown at 56 °C. Thermophilic and hyper-thermophilic bacteria are found in both the eubacteria and the archaea. Most hyperthermophilic archaea are associated with waters or soils which are heated by geothermal activity and which contain sulphides and elemental sulphur. Reactions involving sulphur are an important part of the metabolism of most of these organisms.
Continue reading here:Ffffffffffffff uuuuuuuuuuuu
Was this article helpful?
Recommended Programs

Readers' Questions
-
Mewael2 months ago
- Reply