易感宿主
致命的病原体
图三分之一的植物病害的三角形,说明主机之间的关系,病原体,和环境必要的疾病发生。
有利的环境
图三分之一的植物病害的三角形,说明主机之间的关系,病原体,和环境必要的疾病发生。
个人遇到的伴侣。例如,局部腥黑穗病的影响病原体需要遇到两个繁殖交配类型之间发生,然而其propa-gules风力分散,使得人与人之间的接触不同的交配类型不可能当人口很小。著产生的效应可能有助于解释为什么这个物种还没有更多的入侵,自从遇到交配类型之间时将更有可能被风吹到更大的区域(Garrett和鲍登,2002)。物种体验,这种Allee效应与疾病nonconducive环境条件进一步减少感染的机会。因此,如果气候条件更有利于疾病的病原体是释放著的约束效应,病原体的数量可能会增加比预期快得多。
典型的“复利”发展植物病害流行的病原体和几代人每赛季还可以导致重要阈值的结构。感染水平通常可以增加数量级向本赛季结束。如果生长季节的长度增加,区域生产特定的作物物种可能扩大随着时间的推移,随着时间越长季长度允许大幅增加病原体种群生长季节的末尾。这些人也可能会降低收益率在那个赛季,作为大剂来源即将来临的种植季节。如此高的地区培养液加载可能产生积极的反馈循环,呈现本地应用程序的一些管理技巧那么有用。例如,本地应用程序技术,如卫生(删除受感染的植物材料),使用品种混合物,并使用基于降低培养液的抗病性生产所有依赖,至少在某种程度上,在控制的能力
152年全球气候变化和raybet雷竞技最新极端天气事件
当地培养液负载。如果区域培养液负荷过高,这些方法的贡献将会减少(Garrett et al .,在修订)。同样,在荒地系统中,植物多样性可能提供基线管理的植物病害的赏识,但可能会减少如果系统成为饱和与培养液。相反,如果季节变短或气候条件在生长季节变得不那么重要的地方疾病有利,更大的好处可能会获得一些管理技巧。
病原体范围变化可能发生随着气候条件的变化,使感染和过冬或oversummering新领域。气候变化的影响可能与其他现象,如引入新的病原体种类或病原体的基因型。例如,过冬,推动了马铃薯晚疫病病原的不同的交配类型的存在,使得性生产的顽强合子和潜在的适应通过生产新基因型(Widmark et al ., 2007)。温和的冬天和引入新的交配类型可能大大增加问题这种病原体。范围变化和病原体的介绍也将导致新遇到病原体物种之间杂交产生的潜在的新的病原体(火盆,2001)。同样,新的向量物种的引入或范围的转变可能会使疾病更重要,比如在glassy-winged神枪手的运动以及由此导致的皮尔斯病风险增加的葡萄园(Redak et al ., 2004)。
物候变化和范围的变化对气候变化的反应可能不遵循相同的模式植物宿主和病原体。raybet雷竞技最新一些病原体只能感染特定植物生长阶段或器官,如鲜花。例如,镰刀菌素头枯萎病病原体感染小麦花药或其他植物的器官(白的沙,2004)。花期物候的变化,以应对气候变化可能不匹配等病原体物候学的转变,感染率可能会出人意料地上升或下降。raybet雷竞技最新不同的地理范围变化的模式可能会导致新的宿植病原体组合(帕克和吉尔伯特,2004)。病原体的遗传潜力适应性人群很重要在决定是否产生减少感染将短期或持久的。一般来说,病原体适应的时间表可能是比时间更短植物适应。这将是特别是长寿植物物种在荒地,也为一年生作物物种即使有完整的农业科学家的注意。
政策也可能在一些重要方面与非生物环境交互。随着爱尔兰马铃薯饥荒,另一个戏剧性的例子是在美国中部尘暴区。这个地区政策,支持广泛的土地耕作正值气候条件支持风蚀的土壤。要么单独因素可能造成问题,但两者的结合导致了毁灭性的条件。生物和社会因素的相互作用也可能导致气候变化的放大效应。raybet雷竞技最新例如,如果
raybet雷竞技最新气候、生态、疾病和感染个案我们153
植物病原体故意引入(弗莱彻和堆栈,2007;马登Wheelis, 2003),恐怖主义使用它们可能会寻找最有利的环境条件,建立在脆弱的种植制度。当地的地区食品安全密切与当地粮食生产将特别容易受到作物疾病的压力的变化。然而社会在这些地区也可能倾向于依靠作物物种不太好得到研究和发展的支持。这些“孤儿作物”,如小米、藜麦、木薯、画眉草,需要更多的研究支持缓冲他们是重要的社会的脆弱性(Nelson et al ., 2004)。
应对气候变化raybet雷竞技最新
好消息植物病害管理策略的制定在气候变化条件下需要做的是相同的或没有气候变化。raybet雷竞技最新即使没有气候参数的长期趋势,气候变化从一季到下一季,每年,地区需要知识和工具为适应不同的场景。然而,潜在的气候变量的新组合,以及潜在的相互作用和快速变化的条件下,强化了需要研究和植物病害的应对政策风险(麻省et al ., 1999;加勒特et al ., 2006)。研究直接明确地理解系统应对气候变化的复杂性是必要的。raybet雷竞技最新
机械的理解植物和病原应对气候变化将基于特征适应当前的人口和他们的潜力。raybet雷竞技最新新的基因组工具能够描述基因表达和基因型多样性更容易在荒地和农业植物群落。这些工具可以应用与其他“组学”的方法将反应基因的表达(转录组),lipidomics,代谢组学更全面适应潜力的机械的理解。这些方法将应用于多因素研究气候变化的影响,所以温度变化之间的相互作用的影响,降水、二氧化碳,和其他环境因素可以被理解,以及潜在的适应。raybet雷竞技最新
工具研究病原体人口和社会结构、基因表达和其他反应正在迅速发展。测序技术的进步使微生物群落的常规描述可行(Riesenfeld et al ., 2004;罗斯切et al ., 2007),最终将使它便宜。微阵列,如GeoChip(他et al ., 2007),目的是研究土壤微生物的基因功能。需要新的微阵列研究微生物的存在和表达基因与植物病害。这将是重要的收集基线微生物群落结构和功能的信息很快,这样的变化可以研究新的气候条件下微生物群落。实验比较反应的微生物com -
154年全球气候变化和raybet雷竞技最新极端天气事件
在刚才新环境也将重要的(例如,沃尔德罗普和费尔斯通,2006)。无疑有很多形式的疾病suppressiveness提供的农业土壤中微生物群落提供福利和规范疾病自然系统和目前被低估了。
研究阐明宿主景观结构的影响将有助于改善策略和必要的研究区域的变化,大陆和全球尺度。当前区域分析气候对疾病风险的影响往往是疾病风险计算个人“像素”,重要的发展中第一个近似估计的风险。这种模型的下一个阶段将是把社区提高风险估计,风险对于任何给定的位置会增加接近高风险地区。最后,区域和全球模型需要将病原体的进化。制定和参数化这些模型需要流行病学的理论和实验进展。例如,更好的数据和模型相关病原体和宿主传播,目前的种内差异水平,选择不同的气候变化情况下的强度是必要的。raybet雷竞技最新
长期地域代表性疾病发生的记录和病原体和宿主的分布非常罕见,尽管其重要性理解流行病学和流行趋势(联合工作组和Pautasso, 2008)。全球网络支持流行的分析是必要的。进展这一目标到位;例如,美国发达国家植物诊断网络,以促进数据收集和分析(堆栈和弗莱彻,2007)。是最有效的,这个网络最终需要与在其他国家可比国家网络。雷竞技手机版app是美国的优势来帮助其他国家建立这样的网络来收集和分析数据,以便我们都能受益于更完整的信息。雷竞技手机版app修改农业管理的使用模型预测被证明是有用的在世界的许多地方,包括应用程序资源贫乏的农民基于气候预测在津巴布韦(Patt et al ., 2005)。raybet雷竞技最新
最重要的一个投资我们可以保护,特征,发展策略优化植物遗传资源的使用。在荒地系统中,保护是必要的增加机会足以包括植物种群个体的基因型适应新的气候情况。raybet雷竞技最新在农业系统中,保护作物物种的多样性及其野生亲缘是必要的增加阻力所需基因的可能性,新的生物和宽容非生物压力维护(Johnson, 2008)。原位保护允许自然选择继续作用于这些物种。非原位保护是一个有用的备份策略和简化登记入册的一些分析。国际网络保护作物遗传多样性,如机构在国际协商小组农业研究(CGIAR)是至关重要的,以确保保护和分析登记入册。当前可用的资金等项目相比很低他们的任务的重要性。而投资
raybet雷竞技最新气候、生态、疾病和感染个案我们155
斯瓦尔巴全球种子库提供一个最后贷款人,活跃的植物资源调查是必要的。
最终我们最好的农业应对气候变化的挑战将是发展多样化,灵活、有弹性的农业系统,可以更容易适应新的气候raybet雷竞技最新条件。这些系统必须包括准备和资金充足的农业科学家在全球范围内积极开发新策略。在荒地系统中,替换植物物种或基因型可能是一个不那么有吸引力的选择。自入侵病原体可以有最重要的影响,有可能加剧气候变化的影响,政策更好地减少外来病原体的传播很重要(安德森et al ., 2004;raybet雷竞技最新体细胞杂种et al ., 2006)。
确认
我很欣赏价值的评论论坛成员和员工微生物威胁,r·鲍登·d·罗滕伯格,p·加芬克尔。也是高兴承认支持由美国国家科学基金会(NSF)通过授予deb - 0516046和NSF格兰特ef - 0525712作为联合的一部分NSF-National卫生研究院(NIH)生态传染病项目;由美国国际开发署(USAID)的国际研究、教育、开发(OIRED)的弗吉尼亚理工大学可持续农业和自然资源管理(SANREM)合作研究支持计划(CRSP)奖。epp - a - 00 - 04 - 00013 - 00和害虫综合管理(IPM) CRSP奖。epp - - 00 - 04 - 00016 - 00;贡献和堪萨斯州实验站(08 - 308 b)。
raybet雷竞技最新气候变化和传染病:ARCTIC12对人群的影响
艾伦·j·帕金森Ph.D.13疾病控制和预防中心
作品简介:北极环境
极地附近的区域被定义为该地区纬度延伸超过60,边界北冰洋,和包括所有的八个国家的北部地区:美国(阿拉斯加)、加拿大、格陵兰、冰岛、挪威、
12本报告中的发现和结论的作者,不一定代表官方立场的疾病控制和预防中心。
13北极调查计划,分工新兴感染和监视服务,国家中心准备检测和控制传染病,安克雷奇,正义与发展党。
156年全球气候变化和raybet雷竞技最新极端天气事件
芬兰、瑞典和俄罗斯联邦2 - 20(见图)。的raybet雷竞技最新北极的气候从严寒干旱无人居住的地区不同地理位置毗邻沿海温带森林农业地区。大约有400万人生活在北极,几乎有一半居住在俄罗斯联邦的北部地区。北极和人民被回归线地区生活在社会和物理环境之间有着本质的不同的更多的南方居住。这些数量是由不同比例的原住民和nonindigenous人民(Stephansson北极研究所,2004;参见图2-21)。
加拿大北部的土著人群(魁北克省北部,西北地区,育空,努和拉布拉多),阿拉斯加、格陵兰岛通常位于偏远地区的小社区。他们几乎没有经济基础设施和依赖生存打猎、钓鱼、和采集食物的很大一部分他们的饮食。在这些偏远地区,公共卫生和急性护理系统的访问通常是边际和糟糕的支持。预期寿命原住民的阿拉斯加,加拿大北部,格陵兰岛和低于一般人群的美国,加拿大和北欧国家(年轻,2008)。雷竞技手机版app同样的婴儿道德这些人群的土著段高于可比国家人口。心脏病和癌症死亡率,一旦低得多的土著人群中美国,加拿大和北欧国家,现在类似于各自国家的利率。雷竞技手机版app阿拉斯加土著人群、加拿大和格陵兰儿童意外伤害有较高的死亡率和自杀。北极原住民的其他健康问题包括某些传染病的高发,如乙型肝炎、幽门螺杆菌、呼吸道合胞体病毒(RSV)感染的婴儿,和性传播疾病,以及健康的影响与对环境污染物的暴露,经济快速变化和现代化,和气候变化(Bjerregaard et al ., 2004)。raybet雷竞技最新
raybet雷竞技最新北极气候变化和环境
北极,像大多数世界其他地区一样,温暖大大超过20世纪,主要是最近几十年。北极气候模型项raybet雷竞技最新目持续变暖3-5oC意味着增加2100。冬天会温暖的夏天多,年平均降雨量预计将增加,预计持续融化的土地和海冰增加河道流量,导致海平面上升。这些变化将是伴随着更大的整体气候变化和极端天气事件的增加(北极理事会,2005)。raybet雷竞技最新
迅速在北极变暖已经带来了巨大的生态和社会经济的影响,其中许多由于永久冻土融化,洪水、风暴潮带来的和海岸线侵蚀和
raybet雷竞技最新气候、生态、疾病和感染个案我们157
raybet雷竞技最新气候、生态、疾病和感染个案我们157
-
 with permission from W. K. Dallmann and the International Journal of Circumpolar Health. Copyright 2008.")
- 图2 - 20显示行政辖区极地附近的地区。资料来源:由w . k . Dallmann地图。从年轻(2008)转载许可w . k . Dallmann和国际环极健康杂志》上。2008年版权。
 with permission from W. K. Dallmann and the International Journal of Circumpolar Health. Copyright 2008.")
失去保护的海冰。在许多社区,建立基础设施支持永久冻土。失去这名冻土地基将导致损坏,进水系统和管道,并可能导致污染的社区水的供应。此外,损失的基础支持访问道路、栈道、水储罐和污水处理设施将使配水和废水处理系统瘫痪。几个村庄已经面临搬迁,因为村住房,水系统和基础设施被破坏(沃伦et al ., 2005)。
快速的变暖导致了北极海冰每年的损失。2007年9月11日,北极海冰覆盖面积达到最低程度上记录
158年全球气候变化和raybet雷竞技最新极端天气事件
158年全球气候变化和raybet雷竞技最新极端天气事件
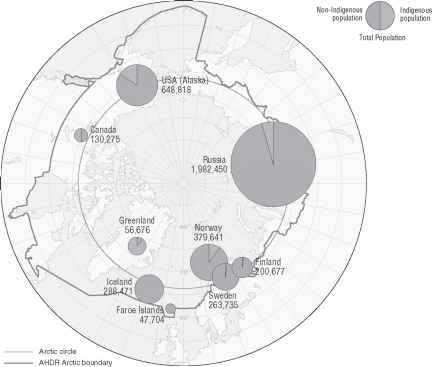
图2-21环极地区展示土著和nonindigenous人口分布。
来源:转载来自Stefansson北极研究所(2004)从w . k . Dallmann许可,挪威极地研究所Stefansson北极研究所。2004年版权。
图2-21环极地区展示土著和nonindigenous人口分布。
来源:转载来自Stefansson北极研究所(2004)同意w . k . Dallmann挪威极地研究所和Stefansson北极研究所。2004年版权。
观察开始于1970年代,超过最悲观的预测模型无冰的北极到2050年(Richter-Menge et al ., 2008;图2-22)。这种戏剧性的海冰的减少会对海洋生态系统产生广泛的影响,沿海气候,人类定居点,生存活动。raybet雷竞技最新第一次年度海冰的减少了无冰的西北航道,从拉布拉多在加拿大北部的北极群岛北部,白令海峡,并几乎完全清除东北之行,从白令海峡沿着北部海岸的俄罗斯联邦挪威(参见图2-23)。路线代表时间,省油捷径太平洋与大西洋之间的要道和将增加海洋运输和获得丰富的石油、天然气和矿藏一旦无法勘探和开发。
raybet雷竞技最新气候、生态、疾病和感染个案我们159
raybet雷竞技最新气候、生态、疾病和感染个案我们159

图2-22北极冰帽,2001年9月2007年9月(上)和(下)。
来源:美国国家航空航天局,印刷Borgerson (2008)。
图2-22北极冰帽,2001年9月2007年9月(上)和(下)。
来源:美国国家航空航天局,印刷Borgerson (2008)。
这样的访问将带来很多好处以及风险一旦孤立北极社区。新海岸警卫队或建设军事基地和其他工业企业将给当地居民带来就业机会,但也会影响人口分布、动态、文化和当地的环境。旅游很可能会增加。公共部门和政府服务将增加新兴经济体的支持。这些事件将大大
160年全球气候变化和raybet雷竞技最新极端天气事件
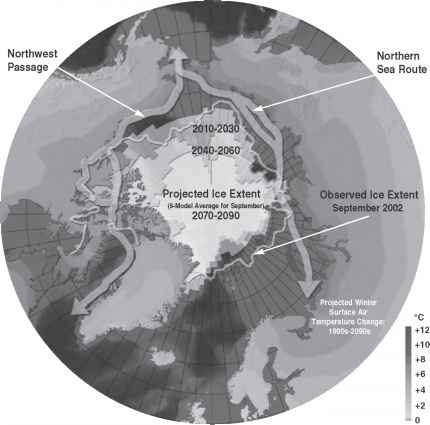
图2-23提出西北和东北航道通过北冰洋加入大西洋和太平洋。
来源:地图的c . Grabhorn重印适配器(2004)剑桥大学出版社的许可和c . Grabhorn。
图2-23提出西北和东北航道通过北冰洋加入大西洋和太平洋。
来源:地图的c . Grabhorn重印适配器(2004)剑桥大学出版社的许可和c . Grabhorn。
挑战传统的自给自足的生活方式对于许多社区和导致快速和长期的文化变革,这将创建额外的压力已经脆弱人群(Curtis et al ., 2005)。
raybet雷竞技最新气候变化和人类健康
气候变化将带来的直接健康影响环境温度的变化,从户外活动模式raybet雷竞技最新改变的风险,传染病的发病率的变化。随着环境温度增加,
raybet雷竞技最新气候、生态、疾病和感染个案我们161
低体温症的发病率和相关的发病率和死亡率可能会降低。相反高热会增加,尤其是非常年轻和老年人(Nayha, 2005)。然而,由于许多北极地区的平均气温较低,这类事件有很大的可能性为普通人群对公共卫生的影响很低。更重要的是,意外伤害,主要是有关生存狩猎和fishing-already北极居民可能增加死亡率的一个重要原因(北极理事会,2005)。减少河流和海冰厚度,减少冰季节,减少了积雪,和冻土融化将使狩猎和采集更加困难,危险,和更少的成功,从而增加溺水伤害和死亡的风险。
永久冻土融化流失或洪水可以强制搬迁。社区和家庭经历搬迁将不得不适应新的生活方式,可能面临失业,必须整合和创造新的社会关系。搬迁也可能导致快速和长期的文化变革和传统文化丧失,这将增加个人和社会压力,导致心理和行为健康挑战(Hess et al .,在出版社)。
raybet雷竞技最新气候变化已经对食品安全构成严重威胁的许多北极社区,因为他们的生存依赖传统的自给自足的狩猎和捕鱼。人口的海洋和陆地哺乳动物、鱼类和水禽可能减少或取代通过改变栖息地和迁徙模式,进一步减少传统的食品供应。发布环境污染物的大气和海冰融化的冰川和可能增加这些污染物的水平进入食物链,使传统食品不太可取的(同理,2003)。减少传统食品供应将迫使原住民社区越来越依赖非传统和西方食物通常不太健康。这很可能会导致增加率与加工食品相关的现代疾病,如肥胖、糖尿病、心血管疾病和食源性传染病的爆发与进口新鲜和加工食品(Bjerregaard et al ., 2004;奥尔et al ., 1994)。
许多开始系统对气候变化特别敏感。raybet雷竞技最新特定阶段的生命周期的许多寄生虫可能极大地受温度的影响。例如,小的增加温度可以大大增加肺的传播蠕虫和肌肉致病性野生动物作为食物来源很重要对于许多北部社区(Hoberg et al ., 2008)。
raybet雷竞技最新在北极气候变化和传染病
众所周知,raybet雷竞技最新气候和天气影响分布和许多病媒传播疾病的风险,如疟疾、裂谷热、瘟疫,登革热在世界的热带地区。天气也会影响食源性和水源性疾病的分布和新兴传染病,如西尼罗河病毒,
162年全球气候变化和raybet雷竞技最新极端天气事件
汉坦病毒,埃博拉病毒出血热(Haines et al ., 2006)。少即是已知的关于气候变化的影响和传染病的风险和分布在北极地区。raybet雷竞技最新众所周知,北极人口历史悠久的流行和流行性传染病(帕金森et al ., 2008)。然而,通过引入抗菌药物,疫苗,和公共卫生系统,由于传染病发病率和死亡率大大降低。尽管有这些进步,高侵袭性疾病引起的肺炎链球菌,流感嗜血杆菌、结核分枝杆菌存在(Bruce et al ., 2008 a, b;克里斯腾森et al ., 2004;Dawar et al ., 2002;Degani et al ., 2008;Gessner et al ., 1998;迈耶et al ., 2008; Netesov and Conrad, 2001; Nguyen et al., 2003; Singleton et al., 2006; S0borg et al., 2001). Sharp seasonal epidemics of viral respiratory infections also commonly occur (Bulkow et al., 2002; Karron et al., 1999; Van Caeseele et al., 2001). The overuse of antimicrobial drugs in some regions has led to the emergence of multidrug-resistant S. pneumoniae, Helicobacter pylori, and methicillin-resistant Staphylococcus aureus (Baggett et al., 2003, 2004; McMahon et al., 2007; Rudolph et al., 1999, 2000).
气候的影响在这些现有的传染病raybet雷竞技最新的发病率是未知的挑战。然而在许多北极地区,住房拥挤,卫生条件不足已经传染性疾病传播的重要决定因素。北方寒冷的气候使人在室内放大家庭聚集raybet雷竞技最新的影响,吸烟,和通风不足。拥挤的生活条件提高人际传播的传染病和呼吸道和胃肠疾病的传播和皮肤感染。许多家庭在北极社区缺乏基本的卫生设施服务(例如,抽水马桶,淋浴或洗澡,厨房水槽)。提供这些服务是困难的在偏远村庄里孤立小种群生活在一个严酷的寒冷气候。raybet雷竞技最新阿拉斯加西部的最近的一项研究表明两到四倍住院率小于3岁的儿童肺炎、流感和童年RSV感染在村子里大多数的家庭内部没有自来水,而村子里大多数的家庭内部自来水服务。同样,门诊和住院治疗皮肤感染金黄色葡萄球菌感染在所有年龄段的人是高的村庄没有内部自来水服务相比的村庄没有水服务(轩尼诗et al ., 2008)。损害公共卫生基础设施由冻土融化或洪水可能因此导致增加利率住院儿童的呼吸道感染,以及增加率引起的皮肤感染和腹泻疾病的细菌,病毒,寄生虫病原体。
还有一些独特的传染病北极土著人口和生活方式,在北极变暖可能会增加。例如,许多北极居民依靠自给狩猎、捕鱼、采集食物,在一个可预测的环境raybet雷竞技最新储存食物。储存食物的方法通常包括地面用电吹风的鱼和肉在环境温度,低于地面
raybet雷竞技最新气候、生态、疾病和感染个案我们163
冷藏或附近的冻土,发酵。气候的变化会防止干燥的raybet雷竞技最新鱼或肉,导致腐败。同样,冻土的损失可能会导致腐败的食物储存在地下。食源性中毒暴发发生零星的社区在美国,加拿大北极,格陵兰岛和准备是由摄入不当引起的传统发酵食品(疾病预防控制中心,2001;•et al ., 1997;Sobel et al ., 2004;S0rensen et al ., 1993;温赖特et al ., 1988)。因为肉毒梭状芽胞杆菌的孢子萌发和毒素产量将出现在温度大于4°C,温暖的环境温度可能与气候变化有关可能导致这些地区的食物中毒率增加。raybet雷竞技最新岁的初步研究表明,发酵海豹肉的挑战与c在4摄氏度以上的高温导致肉毒毒素生产(Leclair et al ., 2004)。
由弧菌引起的胃肠炎暴发parahaemolyticus一直与食用生的或未充分煮熟的贝类来自海水的温度高于15度。最北端的爆发发生在2004年之前,在1997年不列颠哥伦比亚省北部。然而,2004年7月,食品中引起的胃肠炎暴发相关拮抗记录在游轮乘客食用生蚝在威廉王子湾牡蛎农场视察时,阿拉斯加(麦克劳克林et al ., 2005)。疫情调查记录增加每年0.21摄氏度水温自1997年以来,7 - 8月,报道说,2004年是第一年牡蛎农场水温超过7月15摄氏度。这个事件提供了直接证据的上升之间的联系海水温度和发病。
气温升高可能会使感染宿主动物物种生存的冬天在更大的数字,增加人口,扩大范围的居所,因此增加了机会通过感染人类。例如,温和的天气和更少的积雪可能造成了一个大爆发的2007年在瑞典北部Puumala病毒感染。在银行田鼠Puumala病毒流行,在人类导致肾综合症出血热(佩特森et al ., 2008)。类似疫情在俄罗斯联邦(Revich, 2008)。北方气候raybet雷竞技最新的扩张在阿拉斯加和加拿大北部针叶林带青睐海狸的稳定向北推进,扩展的范围贾第虫属lamblia,海狸的寄生虫感染可以感染其他哺乳动物,包括人类使用未经处理的地表水(北极理事会,2005)。同样,北极地区的气温升高被回归线地区可以支持狐狸的地理范围和数量的扩张和田鼠,常见的运营商Echinococ-cus multilocularis,肺泡棘球绦虫引起的人类(霍尔特嗤之以鼻et al ., 2005)。瑞士肺泡棘球绦虫的患病率上升随着福克斯人口增加的规模和地理范围扩大到城市地区(Schweiger et al ., 2007)。肺泡棘球绦虫是常见的两种
164年全球气候变化和raybet雷竞技最新极端天气事件
地区西北部的阿拉斯加1997年之前。疾病在人类与狗接触;然而,在住房和改善狗很多管理在很大程度上消除dog-to-human传输在阿拉斯加。这可能不是这样,然而,在北极的其他部分人类感染棘球绦虫granulosis,和大肠multilocularis仍报道,尤其是在协会与社区依赖驯鹿放牧和狗使用(Castrodale et al ., 2002;Rausch, 2003)。
raybet雷竞技最新气候变化也影响动物宿主和蚊子的密度和分布向量,这可能导致人类疾病的增加或改变疾病的地理范围由这些药物引起的。这些变化对人类疾病发病率的影响还没有被充分评估,但显然是潜在的气候变化将某些媒介传播的地理分布和其他人畜共患疾病。raybet雷竞技最新例如,西尼罗河病毒在1999年进入美国,在随后的年感染人类,马,蚊子,和鸟类种群在美国和北至北部马尼托巴省,加拿大(帕金森和巴特勒,2005)。在俄罗斯联邦受感染的鸟类和人类已发现北地区的新西伯利亚(Revich, 2008)。虽然有,但目前,气候之间的关系信息不足和西尼罗河病毒的传播,许多因素可能有助于其进一步向北迁移。raybet雷竞技最新温和的冬天可能会忙冬季生存受感染的库蚊种虫害蚊子,西尼罗河病毒的主要载体,自1970年代以来已经迁移到北艾伯特王子,在加拿大萨斯喀彻温省。长,炎热的夏季增加受感染蚊子的季节导致更高的数字传播和人类接触的机会。raybet雷竞技最新气候变化可能改变其他的疾病生态学和迁移模式水库如鸟。这些因素可能影响疾病发病率和导致其他节肢动物病媒传播疾病的范围的扩张。
大量的蚊子传播的病毒引起人类疾病传播在美国北极和俄罗斯联邦北部地区(沃尔特斯et al ., 1999)。詹姆斯敦峡谷和北美野兔病毒被认为是新兴的公共卫生威胁在美国,加拿大,俄罗斯联邦,引起类似流感的症状和中枢神经系统疾病,如无菌性脑膜炎和脑炎(沃尔特斯et al ., 1999)。辛德毕斯病毒也在北欧循环。这种病毒是由候鸟北和放大。在夏末,ornithophilic蚊子把病毒传给人类导致芬兰北部Pogosta疾病的流行,一种疾病表现为皮疹和关节炎(Kurkela et al ., 2008)。在瑞典,蜱传脑炎(此种)的发病率大大增加自1980年代中期(林格伦和Gustafson, 2001)。增加对应于一个温和的冬天和春天的早期发病的趋势,导致增加蜱虫人口(Ixodes蓖麻),携带病毒负责此种和其他潜在的病原体(Skarphedinsson et al ., 2005)。同样在北-
raybet雷竞技最新气候、生态、疾病和感染个案我们165
加拿大东部,气候变化预计将导致向raybet雷竞技最新北转移的范围Ixodes scapularis,蜱虫,携带,伯氏疏螺旋体莱姆病的病原体。目前北部Ix的极限。scapularis安大略省南部包括伊利湖的海岸线和新斯科舍省的南部海岸。一些temperature-based模型显示潜在的Ix的向北扩张。scapularis高于60纬度和到2080年西北地区(奥格登et al ., 2005)。然而,应该指出的是,蜱虫分布受到其他因素的影响如栖息地适宜性和传播模式可以影响这些预测的准确性。是否在人类疾病是由于这些气候改变引发的改变向量范围取决于许多因素,如土地利用实践,人类行为(郊区发展在树木繁raybet雷竞技最新茂的地区,户外娱乐活动,使用驱虫剂、等等),人口密度,和充足的公共卫生基础设施。
应对气候变化在北极raybet雷竞技最新
1992年,国际移民组织发表了一份报告《新兴感染:微生物在美国健康威胁。这份报告发现了重大的公共卫生挑战医学界主要相关检测和管理传染病疫情和监测流行传染病的流行。它激发了民族运动重振美国的公共卫生系统应对艾滋病毒/艾滋病的流行,新疾病的出现,旧的疾病死灰复燃,抗菌素耐药性的持续进化。在随后的报告中,国际移民组织提供了一个评估公共卫生系统的能力应对新兴威胁并提出建议,以解决传染病对人类健康威胁(国际移民组织,2003)。
因为气候变化预计raybet雷竞技最新将加剧许多因素的传染病出现再度出现,2003国际移民组织的报告的建议可以应用到新兴传染病的预防和控制气候变化带来的威胁。公共卫生应对气候变化的框架在美国最近提出(Frumkin et al ., 2008raybet雷竞技最新;赫斯et al .,在出版社)。框架强调了需要利用和提高现有基本公共卫生服务和改善政府机构之间的协调工作(联邦、州和地方),学术界、私营部门和非政府组织。
这个框架应用到北极地区需要加强公共卫生能力监控疾病会有大量公共卫生的影响,包括儿童呼吸系统疾病、皮肤感染和腹泻疾病,特别是社区卫生系统失败。监控特定的媒介传播疾病,如西尼罗河病毒,莱姆病,此种,应优先在焦点区域的边缘地区同时支持动物和昆虫,和气候变化可能促进地理raybet雷竞技最新
166年全球气候变化和raybet雷竞技最新极端天气事件
扩张的向量。因为北极地区人口相对较小,在大面积分布广泛,针对性检测重要的新兴趋势与气候相关的传染病可能会被推迟。raybet雷竞技最新这个困难可以克服连接区域监控系统一起共享标准化信息对气候敏感的目的共同关心的传染病。raybet雷竞技最新应努力协调法定传染病注册、临床监测实验室方法,定义允许跨行政区域内类似的疾病报告和分析。这类网络的一个例子是国际极地附近的新兴传染病监测系统。医院和公共卫生实验室这个网络链接在一起的目的监测入侵细菌疾病和肺结核在北极的人群(帕金森et al ., 2008)。
公共卫生能力应增强应对食源性暴发传染病(如肉毒中毒,兰伯氏贾第虫引起的胃肠炎或振捣parahaemolyticus)。公共卫生研究需要确定潜在的气候敏感的基线流行传染病(如西尼罗河病毒,,伯氏疏螺旋体布鲁氏菌spp。棘球绦虫spp,弓形虫spp)。在这两个地区的人类和动物宿主出现可能raybet雷竞技最新的预期。这样的研究可以用来积累更多的证据的影响气候变化对传染病或天气出现,指导早期检测和公共卫生干预策略,并提供科学支持气候变化对公共卫生的行为。raybet雷竞技最新的环极协调研究工作不仅是重要的研究协议,协调实验室方法,数据收集工具,和数据分析,还稀缺资源所产生的影响最大化和最小化研究受影响社区的影响。可以通过现有的国际合作,促进协调等北极理事会14国际环极健康联盟15和环极健康Researchers.16的新成立的国际网络
在北极的挑战,然而,将确保足够的公共卫生能力允许检测疾病暴发和传染病监测趋势最有可能受到气候的影响。raybet雷竞技最新地处偏远的许多社区临床和公共卫生设施,和恶劣的天气条件适当的北极地区,通常排除标本和流行病学数据收集在疫情调查,研究,或持续的监测活动。人员短缺在许多很常见在当地诊所和地区医院已经被日常和紧急护理重点,离开小现有员工的能力帮助公共卫生人员疫情调查,研究,或维护常规监测活动。
14看http://www.arctic-council.org。15看http://www.iuch.org。16日见http://www.inchr.org。
raybet雷竞技最新气候、生态、疾病和感染个案我们167
可能需要额外的资源和培训来确保充足的人员在这些设施,以解决现有的区域差距诊所和医院和公共卫生部门,并确保足够的训练有素的工作人员解决新兴公共气候变化造成的健康影响。raybet雷竞技最新
的一个关键方面的公共卫生应对气候变化在北极地区将以社区为基础的伙伴关系的形成与部落政府社区识raybet雷竞技最新别潜在的威胁和发展策略来应对这些威胁。社区最大的风险,应该针对教育推广,和评估现有的或潜在的健康风险,漏洞,参与设计的以社区为基础的监测和干预策略的制定。基本指标的识别、选择和监测气候变化和社区卫生很重要对于任何应对气候变化在社区一级(Furgal, 2005)。raybet雷竞技最新站点或者village-specific指标的选择应该遵循当地的担忧,可能包括的活动,如关键野生动物或昆虫的监测在一个地区,气候变化可能导致新出现的人畜共患疾病或测量的天气(即。raybet雷竞技最新、降水和温度),水质(即。,turbidity, pathogens), and gastrointestinal illness (i.e., clinic visits) in a community. Linking communities across regions and internationally should facilitate the sharing of standard protocols, data collection instruments, and data for analysis. These linkages will be important for the detection of trends over larger geographic regions, should enhance a community's ability to detect changes that impact health, and will allow the development of strategies to minimize the negative health impacts of climate change on Arctic residents in the future.
结论
居民的北极土著人群特别容易受到气候变化,因为他们的亲密关系,依赖,大地,海洋,和自然资源为他们的文化、社会、经济和身体健康。raybet雷竞技最新增加平均环境温度可能导致食源性疾病的增加,如肉毒中毒和肠胃疾病。平均温度的增加也可能影响发病率的人畜共患传染病和arboviral通过改变人口密度和范围的动物宿主和昆虫。公共卫生应对这些新兴微生物威胁应包括加强公共卫生能力对气候敏感的传染病监测与潜在的大型公共健康影响;raybet雷竞技最新提示对传染性疾病暴发的调查,可能与气候变化有关;raybet雷竞技最新和研究气候和传染病之间的关系出现指导早期检测和公共卫生干预措施。raybet雷竞技最新以社区为基础的监测网络的发展与区域和国家公共卫生机构以及环极健康组织将促进方法
168年全球气候变化和raybet雷竞技最新极端天气事件
标准化、数据共享和传染病的检测趋势在更大的地理区域。这种能力是至关重要的发展策略来最小化的负面影响气候变化对北极居民的健康未来。raybet雷竞技最新
引用参考文献概述
Borgerson, s . g . 2008。北极融化,全球变暖的经济和安全意义。
外交87 (2):63 - 77。克雷蒂安,j . P。,A. Anyamba, S. A. Bedno, R. F. Breiman, R. Sang, K. Sergon, A. M. Powers, C. O. Onyango, J. Small, C. J. Tucker, and K. J. Linthicum. 2007.干旱相关chi-kungunya沿东非沿海出现。美国热带医学和卫生杂志》76 (3):405 - 407。
弗里茨,c . L。,D. T. Dennis, M. A. Tipple, G. L. Campbell, C. R. McCance, and D. J. Gubler. 1996. Surveillance for pneumonic plague in the United States during an international emergency: a model for control of imported emerging diseases. Emerging Infectious Diseases 2(1):30-36. IPCC (Intergovernmental Panel on Climate Change). 2007. Climate change 2007: the physical science basis. Contribution of Working Group I to the fourth assessment report of the IPCC. Cambridge, UK: Cambridge University Press. Linthicum, K. J., A. Anyamba, C. J. Tucker, P. W. Kelley, M. F. Myers, and C. J. Peters. 1999. Climate and satellite indicators to forecast Rift Valley fever epidemics in Kenya. Science 285(5426):397-400.
Stenseth: C。,N. I. Samia, H. Viljugrein, K. L. Kausrud, M. Begon, S. Davis, H. Leirs, V. M. Dubyanskiy, J. Esper, V. S. Ageyev, N. L. Klassovskiy, S. B. Pole, and C. Kung-Sik. 2006. Plague dynamics are driven by climate variation. Proceedings of the National Academy of Sciences 103(35):13110-13115.
Colwell引用
Colwell, r . r . 1996。全球气候和传染raybet雷竞技最新病:霍乱范式。科学》274 (5295):2025 - 2031。
Colwell, R R。,and A. Huq. 1994. Vibrios in the environment: viable but nonculturable Vibrio cholerae. In: Vibrio cholerae and cholera: molecular to global perspectives, edited by I. K. Wachsmuth, O. Olsvik, and P. A. Blake. Washington, DC: American Society for Microbiology. Pp. 117-133.
吉尔,我。,V. R. Louis, I. N. Rivera, E. Lipp, A. Huq, C. F. Lanata, D. N. Taylor, E. Russek-Cohen, N. Choopun, R. B. Sack, R. R. Colwell. 2004. Occurrence and distribution of Vibrio cholerae in the coastal environment of Peru. Environmental Microbiology 6(7):699-706.
罗林斯,T。,G. M. Ruiz, and R. R. Colwell. 2007. Association of Vibrio cholerae O1 El Tor and O139 Bengal with the copepods Acartia tonsa and Eurytemora affinis. Applied Environmental Microbiology 73(24):7926-7933.
(世界卫生组织)。2005年。《疫情周报》80 (31):261 - 268,http://www.who.int回答/ 2005 / wer8031.pdf(2008年5月1日通过)。
raybet雷竞技最新气候、生态、疾病和感染个案我们169
克雷蒂安等。引用
研究人员。,K. J. Linthicum, R. Mahoney, C. J. Tucker, and P. W. Kelley. 2002. Mapping potential risk of Rift Valley fever outbreaks in African savannas using植被指数时间序列数据。摄影测量工程和遥感68 (2):137 - 145。
研究人员。,J. P. Chretien, J. Small, C. J. Tucker, and K. J. Linthicum. 2006. Developing global climate anomalies suggest potential disease risks for 2006-2007. International Journal of Health Geographics 5:60.
Bedno, s。,C. O. Onyango, C. Njugana, R. Sang, S. Gaydos, K. Sergon, and R. F. Breiman. 2006. Outbreak of chikungunya in Lamu, Kenya, 2004. Paper presented at the International Conference on Emerging Infectious Diseases, Atlanta, GA.
CDC(疾病控制和预防中心)。1998年。非洲裂谷fever-East, 1997 - 1998。发病率与死亡率周报》47 (13):261 - 264。
克雷蒂安,J, P。,and K. J. Linthicum. 2007. Chikungunya in Europe—what's next? Lancet 370(9602): 1805-1806.
克雷蒂安,j . P。,A. Anyamba, S. A. Bedno, R. F. Breiman, R. Sang, K. Sergon, A. M. Powers, C. O. Onyango, J. Small, C. J. Tucker, and K. J. Linthicum. 2007. Drought-associated chi-kungunya emergence along coastal East Africa. American Journal of Tropical Medicine and Hygiene 76(3):405-407.
粮农组织(粮农组织)。2006年。可能的裂谷热活动在非洲之角。EMPRES的手表。
国际移民组织(医学研究所)。2003年。微生物的健康威胁:出现、检测和响应。华盛顿特区:美国国家科学院出版社。
联合国政府间气候变化专门委员会(Intergovernmentalraybet雷竞技最新 Panel on Climate Change)。2007 a。raybet雷竞技最新2007年气候变化:物理科学基础。工作组的贡献我的政府间气候变化专门委员会第四次评估报告。英国剑桥:剑桥大学出版社。第三章。
推荐- - - - - -。2007年b。气raybet雷竞技最新候改变2007:自然科学基础。工作组的贡献我的政府间气候变化专门委员会第四次评估报告。英国剑桥:剑桥大学出版社。第十章。
Kovats, r S。,M. J. Bouma, S. Hajat, E. Worrall, and A. Haines. 2003. El Niño and health. Lancet 362(9394): 1481-1489.
林西克姆,k . J。,F. G. Davies, C. L. Bailey, and A. Kairo. 1984. Mosquito species encountered in a flooded grassland dambo in Kenya. Mosquito News 44:228-232.
林西克姆,k . J。,A. Anyamba, C. J. Tucker, P. W. Kelley, M. F. Myers, and C. J. Peters. 1999. Climate and satellite indicators to forecast Rift Valley fever epidemics in Kenya. Science 285(5426):397-400.
Mavalankar D。,P. Shastri, and P. Raman. 2007. Chikungunya epidemic in India: a major public-health disaster. Lancet Infectious Disease 7(5):306-307.
彼得斯,c·J。,and K. J. Linthicum. 1994. Rift Valley fever. In Handbook of zoonoses, Second edition, edited by G. B. Beran. Boca Raton, FL: CRC Press, Inc.
Rezza G。,L. Nicoletti, R. Angelini, R. Romi, A. C. Finarelli, M. Panning, P. Cordioli, C. Fortuna, S. Boros, F. Magurano, G. Silvi, P. Angelini, M. Dottori, M. G. Ciufolini, G. C. Majori, and A. Cassone. 2007. Infection with chikungunya virus in Italy: an outbreak in a temperate region. Lancet 370(9602):1840-1846.
拯救儿童联盟。2007年(1月11日)。非洲之角紧急声明,http://www。savethechildren.net/alliance/media/newsdesk/2007 - 01 - 01. - html(2008年3月4日通过)。
Sergon, K。,C. Njuguna, R. Kalani, V. Ofula, C. Onyango, L. S. Konongoi, S. Bedno, H. Burke, A. M. Dumilla, J. Konde, M. K. Njenga, R. Sang, and R. F. Breiman. 2008. Seroprevalence of chikungunya virus (CHIKV) infection on Lamu Island, Kenya, October 2004. American Journal of Tropical Medicine and Hygiene 78(2):333-337.
170年全球气候变化和raybet雷竞技最新极端天气事件
Tsetsarkin, k。,D. L. Vanlandingham, C. E. McGee, and S. Higgs. 2007. A single mutation in Chikungunya virus affects vector specificity and epidemic potential. PLoS Pathogens 3(12):e201.
联合国(United Nations)。2004年。肯尼亚紧急呼吁,http://www.un.org/depts/ocha/cap/kenya.html(2008年3月4日通过)。
——2006年。早期预警系统的全球调查:评估能力,差距,和机会来建立一个全面的全球早期预警系统自然灾害,http://www.unisdr.org/ppew/info-resources/ewc3/Global-Survey-of-Early-Warning-Systems.pdf(2008年3月4日通过)。
美国瓦茨,d . M。,D. S. Burke, B. A. Harrison, R. E. Whitmire, and A. Nisalak. 1987. Effect of temperature on the vector efficiency of Aedes aegypti for dengue 2 virus. American Journal of Tropical Medicine and Hygiene 36(1):143-152.
(世界卫生组织)。2004年。利用气候预测raybet雷竞技最新传染病暴发:复习一下,http://www.who.int/globalchange/publications/oeh0401/en/(2008年3月4日通过)。
——2006年。基孔肯亚病和登革热在西南印度洋,http://www.who.int/csr/
不/ 2006 _03_17 / en /(2008年3月4日通过)。
——2007 a。裂谷热疫情在肯尼亚、索马里和坦桑尼亚联合共和国,
2006年12月- 2007年4月。《疫情周报》82 (20):169 - 178。
——2007 b。危机中的卫生行动。突出了没有140 - 8到2007年1月14日,http://www.who。
int /工厂/ donorinfo /了/ highlights_140_08_14jan2007。pdf(2008年3月4日通过)。
Stenseth的引用
阿克特曼,M。,K. Zurth, G. Morelli, G. Torrea, A. Guiyoule, and E. Carniel. 1999. Yersinia pestis, the cause of plague, is a recently emerged clone of Yersinia pseudotuberculosis. Proceedings of the National Academy of Sciences 96(24):14043-14048.
研究人员。,J. P. Chretien, J. Small, C. J. Tucker, and K. J. Linthicum. 2006. Developing global climate anomalies suggest potential disease risks for 2006-2007. International Journal of Health Geographics 5:60.
Baltazard, M。,Y. Karimi, M. Eftekhari, M. Chamsa, and H. H. Mollaret. 1963. La conservation interépizootique de la peste en foyer invétéré hypothèses de travail. Bulletin de la Société de Pathologie Exotique 56:1230-1241.
本·阿里T。,A. Gershunov, K. L. Gage, T. Snäll, P. Ettestad, K. L. Kausrud, and N. C. Stenseth. 2008. Human plague in U.S.: the importance of regional and local climate. Biology Letters (in review).
布兰科,g . 1956。一个观点非conformiste苏尔勒德de la有害生物传播模式。Revue d 'Hygiene et医学院学习Sociale 4 (6): 532 - 562。
张伯伦:2004。瘟疫,http://www.kcom.edu/faculty/chamberlain/Website/lectures/lecture/鼠疫。htm(2008年7月1日访问)。
科恩,美国K。,Jr. 2002. The Black Death transformed: disease and culture in early Renaissance Europe. London, UK: Edward Arnold Publishers.
戴维斯。,M. Begon, L. De Bruyn, V. S. Ageyev, N. L. Klassovskiy, S. B. Pole, H. Viljugrein, N. C. Stenseth, and H. Leirs. 2004. Predictive thresholds for plague in Kazakhstan. Science 304(5671):736-738.
戴维斯。,H. Leirs, H. Viljugrein, N. C. Stenseth, L. De Bruyn, N. Klassovskiy, V. Ageyev, and M. Begon. 2007. Empirical assessment of a threshold model for sylvatic plague. Journal of the Royal Society Interface 4(15):649-657.
Drancourt, M。,L. Houhamdi, and D. Raoult. 2006. Yersinia pestis as a telluric, human ectoparasite-borne organism. Lancet Infectious Diseases 6(4):234-241.
raybet雷竞技最新气候、生态、疾病和感染个案我们171
Duplantier, j . M。,J. B. Duchemin, S. Chanteau, and E. Carniel. 2005. From the recent lessons of the Malagasy foci towards a global understanding of the factors involved in plague reemergence. Veterinary Research 36(3):437-453.
埃斯珀,J。,S. G. Shiyatov, V. S. Mazepa, R. J. S. Wilson, D. A. Graybill, and G. Funkhouser. 2003. Temperature-sensitive Tien Shan tree ring chronologies show multi-centennial growth trends. Climate Dynamics 21(7/8):8p.
Frigessi,。,M. Holden, C. Marshall, H. Viljugrein, N. C. Stenseth, L. Holden, V. Ageyev, and N. L. Klassovskiy. 2005. Bayesian population dynamics of interacting species: great gerbils and fleas in Kazakhstan. Biometrics 61(1):230-238.
弗里茨,c . L。,D. T. Dennis, M. A. Tipple, G. L. Campbell, C. R. McCance, and D. J. Gubler. 1996. Surveillance for pneumonic plague in the United States during an international emergency: a model for control of imported emerging diseases. Emerging Infectious Diseases 2(1):30-36.
计,k . L。,and M. Y. Kosoy. 2005. Natural history of plague: perspectives from more than a century of research. Annual Review of Entomology 50(1):505-528.
Galimand, M。,A. Guiyoule, G. Gerbaud, B. Rasoamanana, S. Chanteau, E. Carniel, and P. Courvalin. 1997. Multidrug resistance in Yersinia pestis mediated by a transferable plasmid. New England Journal of Medicine 337(10):677-680.
Guiyoule,。,F. Grimont, I. Iteman, P. A. Grimont, M. Lefevre, and E. Carniel. 1994. Plague大流行调查ribotyping鼠疫杆菌菌株。临床微生物学杂志32 (3):634 - 641。
Guiyoule,。,G. Gerbaud, C. Buchrieser, M. Galimand, L. Rahalison, S. Chanteau, P. Courvalin, and E. Carniel. 2001. Transferable plasmid-mediated resistance to streptomycin in a clinical isolate of Yersinia pestis. Emerging Infectious Diseases 7(1):43-48.
大厅,f·G。,G. Collatz, S. Los, E. Brown de Colstoun, and D. Landis, eds. 2005. ISLSCP Initiative II. DVD/CD-ROM.
Hinnebusch b J。>。t·g·施万,罗素和大肠Carniel》2002。高频接合转移抗生素耐药性跳蚤中肠的鼠疫杆菌基因。分子微生物学46 (2):349 - 354。
霍特兹,p . J。,D. H. Molyneux, A. Fenwick, E. Ottesen, S. Ehrlich Sachs, and J. D. Sachs. 2006. Incorporating a rapid-impact package for neglected tropical diseases with programs for HIV/AIDS, tuberculosis, and malaria. PLoS Medicine 3(5):e102.
亨廷顿,t . g . 2006。强化的证据全球水循环:回顾和合成。水文学杂志319 (1 - 4):83 - 95。
Inglesby, t . V。,D. T. Dennis, D. A. Henderson, J. G. Barlett, M. S. Ascher, E. Eitzen, A. D. Fine, A. M. Friedlander, J. Hauer, J. F. Koerner, M. Layton, J. McDade, M. T. Osterholm, T. O'Toole, G. Parker, T. M. Perl, P. K. Russell, M. Schoch-Spana, and K. Tonat. 2000. Plague as a biological weapon. Journal of the American Medical Association 283(17):2281-2290.
联合国政府间气候变化专门委员会(Intergovernmentalraybet雷竞技最新 Panel on Climate Change)。2007年。raybet雷竞技最新2007年气候变化影响、适应和脆弱性。第二工作组的贡献的政府间气候变化专门委员会第四次评估报告。raybet雷竞技最新英国剑桥:剑桥大学出版社。第八章。
Kausrud, K。,H. Viljugrein, A. Frigessi, M. Begon, S. Davis, H. Leirs, V. Dubyanskiy, and N. C. Stenseth. 2007. Climatically driven synchrony of gerbil populations allows large-scale plague outbreaks. Proceedings: Biological Sciences 274(1621):1963-1969.
Kausrud, k . L。,H. Viljugrein, A. Frigessi, M. Begon, S. Davis, H. Leirs, T. Ben Ari, and N. C. Stenseth. 2008. The epidemiological history of plague in Central Asia: a paleoclimatic modelling study Proceedings of the National Academy of Sciences (in review).
柯伊拉腊,j . 2006。瘟疫:疾病、管理和识别行为恐怖主义。传染性疾病诊所北美20(2):八世,273 - 287。
172年全球气候变化和raybet雷竞技最新极端天气事件
Laudisoit,。,H. Leirs, R. H. Makundi, S. Van Dongen, S. Davis, S. Neerinckx, J. Deckers, and R. Libois. 2007. Plague and the human flea, Tanzania. Emerging Infectious Diseases 13(5):687-693.
洛杉矶,S。,G. Collatz, P. Sellers, C. Malmström, N. Pollack, R. Defries, L. Bounoua, M. Parris, C. Tucker, and D. Dazlich. 2000. A global 9-year biophysical land surface data set from NOAA AVHRR data. Journal of Hydrometeorology 1:183-199.
Mudur, g . 1995。印度的肺鼠疫疫情继续挡板。英国医学杂志》311 (7007):706。
公园,年代。,K. S. Chan, H. Viljugrein, L. Nekrassova, B. Suleimenov, V. S. Ageyev, N. L. Klassovskiy, S. B. Pole, and N. C. Stenseth. 2007. Statistical analysis of the dynamics of antibody loss to a disease-causing agent: plague in natural populations of great gerbils as an example. Journal of the Royal Society Interface 4(12):57-64.
Parkhill, J。,B. W. Wren, N. R. Thomson, R. W. Titball, M. T. G. Holden, M. B. Prentice, M. Sebai-hia, K. D. James, C. Churcher, K. L. Mungall, S. Baker, D. Basham, S. D. Bentley, K. Brooks, A. M. Cerdeno-Tarraga, T. Chillingworth, A. Cronin, R. M. Davies, and P. Davis. 2001. Genome sequence of Yersinia pestis, the causative agent of plague. Nature 413(6855):523-527.
Parmenter, R R。,E. P. Yadav, C. A. Parmenter, P. Ettestad, and K. L. Gage. 1999. Incidence of plague associated with increased winter-spring precipitation in New Mexico. American Journal of Tropical Medicine and Hygiene 61 (5):814-821.
Pettorelli, N。,J. O. Vik, A. Mysterud, J.-M. Gaillard, C. J. Tucker, and N. C. Stenseth. 2005. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in Ecology and Evolution 20(9):503-510.
拉乌尔,D。,and G. Aboudharam. 2000. Molecular identification by "suicide PCR" of Yersinia pes-tis as the agent of medieval Black Death. Proceedings of the National Academy of Sciences 97(23): 12800-12803.
赛米亚:我。,K.-S。Chan和n c Stenseth》2007。广义的阈值混合模型分析非正态的非线性时间序列,应用程序在哈萨克斯坦瘟疫。生物统计学94 (1):101 - 118。
施拉格,s . J。,and P. Wiener. 1995. Emerging infectious diseases: what are the relative roles of ecology and evolution? Trends in Ecology and Evolution 10(8):319-324.
斯科特,S。,and C. J. Duncan. 2001. Biology of plagues: evidence from historical populations. Cambridge, UK: Cambridge University Press.
Stenseth: c . 1999。人口周期在田鼠和旅鼠:密度依赖和依赖阶段在一个随机的世界。Oikos 87 (3): 427 - 460。
Stenseth: C。,A. Mysterud, G. Ottersen, J. W. Hurrell, C. Kung-Sik, and M. Lima. 2002. Ecological effects of climate fluctuations. Science 297(5585):1292-1296.
Stenseth: C。,N. I. Samia, H. Viljugrein, K. L. Kausrud, M. Begon, S. Davis, H. Leirs, V. M. Dubyanskiy, J. Esper, V. S. Ageyev, N. L. Klassovskiy, S. B. Pole, and C. Kung-Sik. 2006. Plague dynamics are driven by climate variation. Proceedings of the National Academy of Sciences 103(35):13110-13115.
Stenseth: C。,B. B. Atshabar, M. Begon, S. R. Belmain, E. Bertherat, E. Carniel, K. L. Gage, H. Leirs, and L. Rahalison. 2008. Plague: past, present, and future. PLoS Medicine 5(1):e3.
Treydte, k . S。,G. H. Schleser, G. Helle, D. C. Frank, M. Winiger, G. H. Haug, and J. Esper. 2006. The twentieth century was the wettest period in northern Pakistan over the past millennium. Nature 440(7088):1179-1182.
Twigg, g . 1984。黑死病:一种生物重新评价。英国伦敦:蝙蝠
继续阅读:信息Dmx
这篇文章有用吗?
推荐项目



读者的问题
-
阿尔特5个月前
- 回复