发现Riftia pachyptila共生关系
科学理解的化学合成的共生,很大程度上源于研究独特的深海动物群热液喷口。早期的探索表明,相比常见的看法,深海底并不感冒,食品有限的栖息地,而是包含了繁荣的生态系统本地化在温泉来自大洋中传播中心。首先在加拉帕戈斯裂谷和东太平洋,东太平洋海隆热液喷口被证明支持高浓度的独立生存的微生物和密集的聚合的无脊椎动物,包括vestimentiferan多毛虫Riftia pachyptila(图1;朗斯代尔1977;Grassle 1985;范多佛2000)。科学家首次提出了悬浮颗粒有机物和独立生存的化能自养的细菌被从水中过滤列支持丰富的无脊椎动物种群(朗斯代尔1977;威廉姆森et al . 1979年)。很快但研究显示,成人r . pachyptila缺乏口腔和肠道琼斯(1981),因此无法暂停进食。看来多毛虫营养,因此流量通过排气食物链的能量,而不是大幅依赖内共生细菌化学合成的。
-

- 图1所示。在东太平洋海隆Riftia pachyptila管虫。鳃羽,gill-like器官用于气体和代谢物交换,突出于白色chiti-nous管保护每个虫的身体

最初的证据在r . pachyptila来自微观化能自养的共生和生化分析显示革兰氏阴性细菌内的包装营养体高度的血管器官多毛虫树干(瓦诺et al . 1981年)。额外的分析涉及稳定同位素(劳1981),酶(Felbeck 1981;Renosto et al . 1991年)和生理(费舍尔et al . 1988年)特征充分表明,r . pachyptila氧化的内共生体减少硫化合物(例如,硫化氢)合成ATP用于自养碳固定通过卡尔文循环(即。“thioautotrophy”)。主机多毛虫能够吸收和运输所需的基质thioautotrophy (HS -、O2和CO2),作为回报,接收部分有机物合成的共生者人口(图2)。细菌的主要手段是人口的碳共生的收购,成人多毛虫,鉴于其无法以可吸入颗粒物为食,是完全依赖于其营养的共生体。
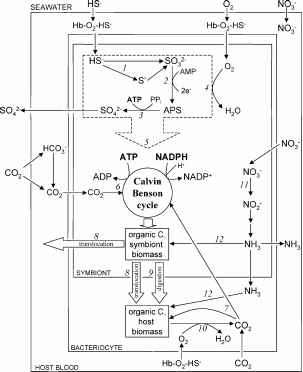
图2所示。提出了模型的代谢之间的共生Riftia pachyptila和化学合成的sulfur-oxidizing细菌。减少硫(主要是HS -)和NO3进入多毛虫血液从环境中通过身份不明的传输机制。二氧化碳和氧气扩散进入。同时血液中,HS -和O2和可逆结合血红蛋白(Hb-O2-HS)运输营养体,这些基质用于共生者硫化物氧化(虚线框)。HS -首先被氧化元素硫(S0)或直接亚硫酸盐(SO32 -;1)。SO32 -氧化硫酸(SO42 -)然后通过APS通过酶通路APS收益还原酶(2)和ATP硫酸化酶(3),产生ATP的磷酸化作用。电子解放硫氧化过程中通过一个电子传递系统,驾驶耗氧量(4)和ATP和NADPH(5)的生产。固定二氧化碳的发生主要是通过ribulose-1 5-bisphosphate加氧酶(二磷酸核酮糖羧化酶)在卡尔文本森周期(6),使用ATP和NADPH产生硫氧化。Anaplerotic通路在主机和共生有机体(7)修复较小数量的二氧化碳。将有机物从共生体通过两主机发生易位简单的营养物质(如氨基酸)发布的细菌(8)和共生有机体的细胞(9)的直接消化。主机耗氧量(10)发生在分解代谢和合成代谢途径的典型。硝酸盐(硝酸盐),占主导地位的共生的氮源,进入通过鼻窦传输机制,减少亚硝酸盐(NO2)的共生体通过同化硝酸还原酶(11)。NO2 -减少通过一个无特征路径产生氨(NH3),用于生物合成共生者和主机(12)。缩写:APS,腺苷5’-phosphosulfate。从趋势Microbiol允许转载(Stewart et al . 2005年)
发现这一专性共生促使调查人员寻找类似的共生,在喷口和其他海洋栖息地(例如,减少沉积物,烃渗漏)。迄今为止,化学合成的细菌被发现与无脊椎动物宿主共生6门以及纤毛虫原生生物(见评论在瓦诺et al . 2005年)。的确,共生菌的存在是一个定义一些类群的特征。例如,所有的成员多毛虫家庭Siboglinidae检查日期,包括vestimentiferan(例如,r . pachyptila)和较小的pogonophoran管虫,包含胞内共生体。这个家庭的成员不仅发生在深海热液喷口(如Oasisia, Ridgeia, Riftia, Tevnia sp。McMullin et al . 2003年)也在寒冷烃渗漏(如Escarpia, Lamellibrachia sp。Sibuet Olu 1998)和泥火山(例如,Oligobrachia;Pimenov et al . 2000;Gebruk et al . 2003年)。这些蠕虫是化学合成的硫的共生体的氧化剂,但methane-oxidizing细菌(氧化菌)被发现在一个宿主物种,pogonophoran多毛虫Siboglinum poseidoni (Schmaljohann和Flugel 1987;见下面章节)。
基于分析16 s rRNA基因序列比较,共生体的通风管虫属于γ变形菌门,一个广泛的细菌分裂,其中包括化学合成的共生体的主机类群多样性(图3,改编自McKiness 2004;瓦诺et al . 2005年)。相比其他化学合成的共生体的γ变形菌门和ε变形菌门的虾和alvinellid episymbionts蠕虫,16 s rRNA基因序列共生者的r . pachyptila集群与其他序列共生体的管虫,包括另一个东太平洋海隆发泄多毛虫,Ridgeia piscesae,和两种渗透管虫,Escarpia spicata和Lamellibrachia一列圆柱(图3)。这个结果支持先前的研究显示,从热液喷口共享一个vestimentiferan管虫,或者非常相似,共生者phylotype (Feldman et al . 1997;劳厄和尼尔森1997;di Meo et al . 2000;纳尔逊和费舍尔2000;McMullin et al . 2003年)。外的“多毛虫集团”Riftia共生有机体似乎最密切相关的化学合成的共生体的热带lucinid蛤(图3)。有趣的是,lucinid蛤和vestimentiferan通风管虫似乎从池中获取他们的共生体的浮游细菌(例如、环境传播,看到格罗斯等人。1998年、2003年和14.4节)。然而,在多大程度上独立生存的共生有机体阶段促进多个主机的入侵(例如,管虫和蛤)在进化的时间仍然是模棱两可的。
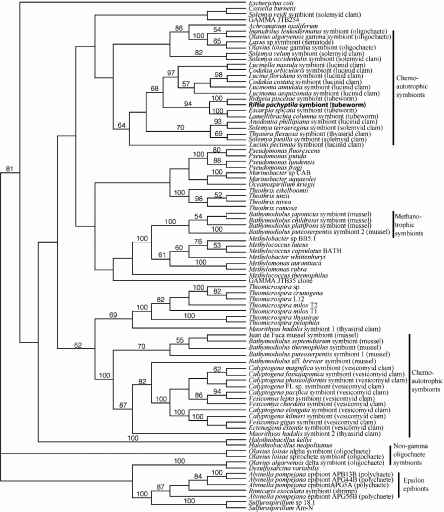
图3所示。系统显示的位置Riftia共生有机体(粗体)相对于其他化学合成的共生体内γ变形菌门,内episymbiontsε变形菌门,和独立生存的细菌。46个树的树是一个严格的共识获得通过吝啬16 s rRNA基因序列的分析(1456个基点)。结果大于50%以上500复制引导分析报告各自的分支。共生体被主机,主机的共同名称分类组括号中列出。改编自McKiness(2004)和瓦诺et al。(2005)
图3所示。系统显示的位置Riftia共生有机体(粗体)相对于其他化学合成的共生体内γ变形菌门,内episymbiontsε变形菌门,和独立生存的细菌。46个树的树是一个严格的共识获得通过吝啬16 s rRNA基因序列的分析(1456个基点)。结果大于50%以上500复制引导分析报告各自的分支。共生体被主机,主机的共同名称分类组括号中列出。改编自McKiness(2004)和瓦诺et al。(2005)
r . pachyptila,至于其他海洋化学合成的昆虫,目前大部分的研究主要集中在澄清的机制,调解共生体的环境采集以及传输的碳,营养,和硫化物共生。这些过程对于理解至关重要的生态和进化主机和共生有机体。但说明这些机制的第一需要知识的分布和生理生化的环境,决定了r . pachyptila。
继续阅读:解剖学和超微结构
这篇文章有用吗?