达尔文Mastotermes及其鞭毛的糖酵解活性
到目前为止,人们一直认为后肠鞭毛虫利用自己的纤维素分解酶生产营养物质,以造福它们的白蚁宿主。在分离细胞中发现的糖酵解活性Mixotrichaparadox在表5.3中编译(Berchtold和König, unpubl。>)。令人惊讶的是,并非所有这些活动都是由鞭毛虫自己产生的,而是由肠道内容物产生的。两种内切葡聚糖酶,Cel I和Cel II,
单毛滴虫属,卡式单毛滴虫,红毛滴虫,凯氏假毛滴虫
人五滴虫,巨型假锥虫
Pentatrichomonoides scroa
毛毛滴虫,阴道毛滴虫,低毛滴虫Trichamäus毛毛滴虫菌株R105毛毛滴虫菌株BUS毛毛滴虫胎
八枝老枝老枝老枝老枝老枝多螺旋体老枝老枝无性克隆Cf5
独立生存的
Monocercomo -
nadidae
Trie ho monad inae
Trichomo-nadinae
P. adamsoni共生体克隆gp 2-2 (Joenina) Metadevescovina extranea Dettotrichonympha nanalD。opercuiata
海雀花
禁忌斯奈德菌无性系3草箭onympha无性系1脆弱阿米巴
——海葵
■假滴管草假滴管
披发虫属麦格纳
柔毛滴虫
De ves covin ida e and
Calonymphidae
过度
mas -
tigea
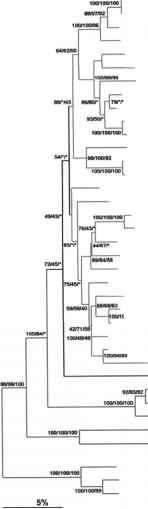
图5.4。副刺属植物的无根系统发育树。SSU rDNA序列的近邻连接分析。Bar表示每100个核苷酸中有5个取代。bootstrap值由三种不同的重建方法计算:距离矩阵,最大parsony和最大似然。星号表示引导值低于40%的节点(Li 2003)。
它们的分子质量约为48kd,是从最原始的澳大利亚白蚁Mastotermes darwiniensis后肠中尚不能培养的共生鞭毛虫中分离出来的(Li et al. 2003)。这些纤维素酶的n端序列与白蚁来源的纤维素酶具有明显的同源性,均属于糖基水解酶家族9。对应的基因不在鞭毛藻的mRNA库中检测到,而是在达尔文Mastotermes darwiniensis的唾液腺中检测到。利用白蚁Mastotermes darwiniensis纤维素酶多克隆抗血清进行western blot分析,在这些鞭毛的粗提物中检测到一种分子质量约为48kd的蛋白质。结果表明,从鞭毛虫营养液泡中分离出的纤维素酶来源于白蚁宿主。这些纤维素酶可能是由达尔文Mastotermes darwiniensis的唾液腺分泌的。在机械研磨过程中对木材颗粒进行研磨白蚁时,纤维素酶附着在木颗粒上或与木颗粒混合,然后附着的纤维素酶或混合物移动到后肠,在那里它们最有可能被鞭毛虫内吞。
表5.3。悖论拌毛糖酵解酶和漆酶活性的测定
酶活性[|U/细胞]
a-L-Arabinosidase 0.3
P-L-Arabinosidase -
P-D-Cellobiosidase 1.2
a-D-Galactosidase 0.2
P-D-Galactosidase -
P-D-Glucosidase 4.0
P-D-Glucuronidase -
a-D-Mannosidase -
P-D-Mannosidase 0.2
P-D-Xylosidase 0.5
169.0纤维素酶
木聚糖酶135.0
2, 2-azinobis-3-ethylbenzthiazolinesulfonic酸
研究表明,对于台湾乳白蚁,这种白蚁的内吞葡聚糖酶仅限于唾液腺、前肠和中肠(Nakashima et al. 2002)。根据我们的工作,在达尔文Mastotermes darwiniensis后肠鞭毛细胞中发现的主要内切葡聚糖酶活性可能来源于白蚁的纤维素酶。研究还发现,达尔文Mastotermes darwiniensis 40%的内切葡聚糖酶活性存在于后肠,而整个后肠的大部分(约84%)纤维素酶活性存在于鞭毛提取物中(Veivers et al. 1982)。这意味着从唾液腺分泌的一定量的白蚁纤维素酶进入后肠并进入鞭毛细胞。它们可能参与鞭毛细胞中纤维素的消化。
采用pcr方法,从鞭毛虫Koruga bonita、Deltotrichonympha nana和D. operculata的微处理细胞核中获得了糖基水解酶家族45的DNA编码纤维素酶。白蚁共生原生生物的纤维素酶序列为系统发育的单系,氨基酸同源性超过84%。推导出的白蚁来源和鞭毛来源的纤维素酶序列由单一的催化结构域组成,缺乏在大多数微生物纤维素酶中发现的纤维素结合结构域(CBD)和间隔序列(Li et al. 2003)。
虽然鞭毛虫内切葡聚糖酶基因在体内也有表达,但SDS-PAGE在营养液泡中未发现明显的鞭毛虫来源纤维素酶活性。western blot检测未检测到鞭毛虫纤维素酶蛋白。这表明鞭毛虫来源的天然内切葡聚糖酶具有很低甚至没有CMC活性,翻译效率也可能很低。在似混毛虫的情况下,没有检测到纤维素酶基因。
可以想象,在2 ~3亿年的过程中,白蚁和最初自由生活的肠道鞭毛虫之间的共生关系相互影响了对方的酶设备,以至于白蚁纤维素酶的过量导致了鞭毛虫自身酶的废弃。后肠缺乏选择压力可能导致纤维素分解酶的低水平翻译、突变和失活,相应的基因仍然表达。无活性酶的产生可能导致相应基因的完全丧失。这意味着共生鞭毛虫正在进入一种没有自身纤维素分解活性的状态,这可能是在含有纤维素的植物存在之前的情况(尽管一些生产纤维素的微生物,如醋酸杆菌,出现得比植物早,但微生物纤维素的数量应该比植物少得多)。目前,澳大利亚原始白蚁Mastotermes darwiniensis的共生肠道鞭毛虫将其内纤维素溶解活性归功于它们的宿主。
继续阅读:整倍体为何保持其外延体
这篇文章有用吗?
读者的问题
-
luam一年前
- 回复