氢化酶体和线粒体遗迹细胞器不断进化而来
最初,参数纺锤剩体,线粒体细胞器残余,氢化酶体几次进化是基于观察氢化酶体和纺锤剩体,分别被发现在一个广泛的,而无关的单细胞生物类群,如
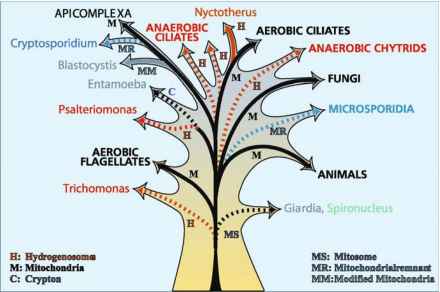
图1所示。插图显示有氧和无氧原生生物之间的系统发育关系(基于各种各样的分子数据)和线粒体的试探性的进化树,一起修改线粒体,线粒体残余,氢化酶体。实线表示的系统发育关系是基于“线粒体基因组的分析。虚线表示organellar基因组的损失。暂定的各种细胞器之间的系统发育关系属于“线粒体”家庭因此基于核基因编码organellar蛋白质的分析。树的这些部分可能被横向基因转移,有缺陷的进化改变目标的各种亚细胞车厢,和有偏见的进化率。有关详细信息,请参阅表1和文本
图1所示。插图显示有氧和无氧原生生物之间的系统发育关系(基于各种各样的分子数据)和线粒体的试探性的进化树,一起修改线粒体,线粒体残余,氢化酶体。实线表示的系统发育关系是基于“线粒体基因组的分析。虚线表示organellar基因组的损失。暂定的各种细胞器之间的系统发育关系属于“线粒体”家庭因此基于核基因编码organellar蛋白质的分析。树的这些部分可能被横向基因转移,有缺陷的进化改变目标的各种亚细胞车厢,和有偏见的进化率。有关详细信息,请参阅表1和文本滴虫,diplomonads,肉足纲(entamoebids)、鞭毛虫,纤毛虫和壶(图1;Biagini et al . 1997;Embley et al . 1997;罗杰·1999)。
此外,厌氧的存在mitochondrion-like细胞器在酵母菌属hominis (Straminopiles)提出了——主要是基于氧化还原敏感的染料(Nasirudeen褐2004)。然而,由于这些细胞器不保留一个基因组,似乎只有常见的诊断特征(i)的存在“mitochondrial-type”chaperonines(克拉克和罗杰1995;Bui et al . 1996年,Germot et al . 1996年),(2)这些细胞器膜是有界的,和(3),在氢化酶体的情况下,这些细胞器产生ATP穆勒(1993)。的系统发育分析HSP (cpn) 60支持不同“线粒体”起源的各种细胞器(Voncken 2001;Voncken et al . 2002;van der Giezen et al . 2003年)。
门 |
订单 |
物种 |
细胞器 |
HSP60 |
氢化酶 |
丙酮酸金属底座。 |
Comp。能源金属底座。 |
细胞器基因组 |
产甲烷内共生体 |
鞭毛纲 |
毛滴虫目 |
毛滴虫(Trichomonadinae) |
氢化酶体 |
HSP60 |
(铁) |
卵圆孔未闭 |
是的 |
没有 |
没有 |
双滴虫目 |
贾第虫属(Giardiinae) |
Crypton |
cytopl.HSP60 |
(Fe) cytopl。 |
卵圆孔未闭 |
没有 |
没有 |
没有 |
|
Spironucleus (Hexamitidae) |
没有确定 |
cytopl.HSP60 |
(铁) |
卵圆孔未闭 |
没有 |
没有 |
没有 |
||
Pelobiontida |
Mastigam oeba (Mastigamoebidae) |
没有确定 |
ND |
(铁) |
卵圆孔未闭 |
ND |
没有 |
没有 |
|
纤毛亚门 |
Armophorida |
Nyctotherus (Clevelandellidae) |
氢化酶体 |
HSP60 |
(Fe) 24 kd + 51 kd |
PDH |
是的 |
是的 |
是的 |
Metopus (Armophoridae) |
氢化酶体 |
ND |
(铁) |
ND |
是的 |
是的 |
是的 |
||
Vestibuliferida |
Dasytricha (Isotrichidae) |
氢化酶体 |
ND |
(铁) |
卵圆孔未闭 |
是的 |
没有 |
没有 |
|
Plagiopylea |
Trimyema (Plagiopylidae) |
氢化酶体 |
ND |
是的 |
PFL |
是的 |
ND |
是的 |
|
Plagiopyla (Plagiopylidae) |
氢化酶体 |
ND |
是的 |
ND |
ND |
ND |
是的 |
||
根足虫类 |
变形目 |
痢疾(Lobosea) |
纺锤剩体 |
HSP60 |
(Fe) cytopl。 |
卵圆孔未闭 |
没有 |
没有 |
没有 |
Percolozoa |
Schizopyrenida |
Psalteriomonas (V ahlkamphiidae) |
氢化酶体 |
ND |
(铁) |
ND |
是的 |
没有 |
是的 |
Alveolata |
Apicomplexa |
Cryptosporidia (Cryptosporidiidae) |
Relict-mitochondrion |
Cpn60 |
NARF |
PNOR |
没有 |
没有 |
没有 |
Stramenopiles |
酵母菌属(Stramenopiles) |
修改mitotochondrion |
cytopl.HSP70 |
没有 |
未知的 |
ND |
是的 |
没有 |
|
小孢子虫目 |
Pansporablastina |
跟踪ipleistophora (P leistophoridae) |
Relict-mitotochondrion |
没有HSP70 |
没有 |
雏形PDH |
没有 |
没有 |
没有 |
Chytridiomycota |
Spizellomycetales |
Piromyces (Neomasticalligales) |
氢化酶体 |
HSP60 |
(铁) |
PFL |
是的 |
没有 |
没有 |
Neocallimastix (Neomasticalligales) |
氢化酶体 |
HSP60 |
(铁) |
PFL |
是的 |
没有 |
没有 |
缩写:;丙酮酸铁氧还蛋白氧化还原酶(卵圆孔未闭);丙酮酸脱氢酶(PDH);丙酮酸甲酸裂解酶(PFL);
热休克蛋白(HSP);Chaperonin (Cpn);丙酮酸NADH氧化还原酶(PNOR);Hydrogenosomal膜蛋白(HMP);
不确定(ND)。
然而,功能和系统发育分析hydrogenosomal ADP / ATP运营商(AAC)透露,迄今为止只有chy-trids和纤毛虫拥有真正的氢化酶体线粒体AAC,集群的线粒体有氧的同系物,mitochondria-bearing亲戚(Voncken 2001;Voncken et al . 2002;Haferkamp et al . 2002;van der Giezen et al . 2002年)。毛滴虫使用替代-潜在pre-mitochondrial aac其hydrogenosomal跨膜运输ATP (Dyall et al . 2000;Tjaden et al . 2004年)。此外,没有证据的存在真正的线粒体aac的纺锤剩体/ cryptons或线粒体遗迹细胞器直到现在。当然,后者不产生ATP,因此,可能不需要mitochondrial-type aac(坠毁et al . 1999, 2003;来自设在et al . 2001;威廉姆斯et al . 2002年)。 Also, the genome projects of Giardia and Entamoeba could not reveal the presence of true mitochondrial AACs, seehttp://www.NCBI.nih.gov。因此,这些观察结果可能主张一个深进化趋异的这些细胞器(兼性厌氧),hydrogen-producing pre-mitochondrial祖先,或者真正的线粒体aac的二次损失在所有这些细胞器(Tjaden et al . 2004年)。在任何情况下,可用的证据强烈支持至少三到四(但可能更多)独立起源的氢化酶体和mi-tochondrial-remnant细胞器细胞器属于“线粒体”家庭——不管这些祖先的细胞器是严格好氧或兼性厌氧(图1;Embley et al . 2003年)。
假设一个有氧健身,线粒体祖先,主机可以依赖的供应与ATP生成线粒体电子传递链的援助。然而,在厌氧条件下,线粒体电子传递链不能使用氧气作为终端电子受体和节能由ATP的生成函数不能满足(2001年Hackstein et al . 1999年)。这样一个细胞面临一个戏剧性的积累减少线粒体等价物,,因此,它必须依靠糖酵解在细胞质中,可以只提供非常有限的ATP。值得注意的是,某些线粒体可以应付缺氧的环境维护某些线粒体功能甚至在这样不利的条件。这些线粒体是能够使用替代环保电子受体(如硝酸盐),或代谢(克雷布斯循环)fu-marate等中间体内源性电子受体允许电子传递链的基本功能甚至在完全没有氧气(Tielens et al . 2002年)。然而在其他真核生物,适应缺氧环境导致的线粒体变性细胞,而不显眼的隔间,相应损失的电子传递链及其节能能力,正如前面所讨论的(Embley和马丁1998;生搬硬套et al . 2000年)。某些线粒体功能,如乙酰辅酶a的生产和keto-acids脂类和碳水化合物的生物合成,分别是维护即使没有一个有效的电子传递链——尽管在另一个亚细胞室(Akhmanova et al。
1998 b;穆勒1998;Hackstein et al . 1999;Henze和马丁2003;Dyall et al . 2004年)。
这篇文章有用吗?