二元信号转导系统
许多细菌,包括诉fischeri、识别和应对自己的环境中使用双组分调节系统(图2,检查存货et al . 2000年)。这些系统是由一个传感器组氨酸激酶
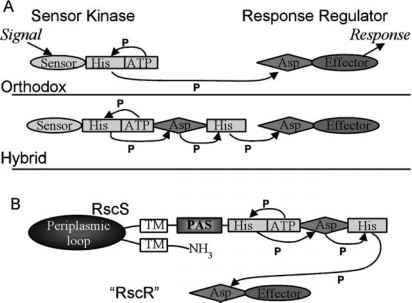
图2所示。双组分调控系统。a的phospho-relay正统(上)和混合(底部)双组分系统。在检测信号,从绑定ATP生成磷酸从守恒的传递给Asp残留直到最后被转移到一个Asp反应调节器,导致反应,改变转录或蛋白质功能。b . rsc激酶是一种混合传感器。rsc的传感器领域是由两个跨膜螺旋(TM),一个大周质的循环和一个不是域
图2所示。双组分调控系统。a的phospho-relay正统(上)和混合(底部)双组分系统。在检测信号,从绑定ATP生成磷酸从守恒的传递给Asp残留直到最后被转移到一个Asp反应调节器,导致反应,改变转录或蛋白质功能。b . rsc激酶是一种混合传感器。rsc的传感器领域是由两个跨膜螺旋(TM),一个大周质的循环,不是域蛋白质识别和传递环境信号(通过自身磷酸化在他的残渣)第二个蛋白质,响应监管机构,(当磷酸化守恒的Asp残渣)进行响应。最频繁,响应由基因表达的改变;另外,蛋白质活动可以变化的结果。
由于环境条件的变化,诉fischeri细胞经历从海水LO,难怪殖民诉fischeri双组分需要监管机构。至少需要两个这样的监管机构的有效起始共生殖民:传感器激酶rsc (Visick和Skoufos 2001)和响应调节器,GacA(2003年惠斯勒和Ruby)。转录监管机构,FlrA展览有限相似反应需要监管机构和起始(2003年米利根和Ruby)也将在这里讨论。
rsc。突变rsc的能力严重降低诉fischeri发起共生殖民:大多数动物保持uncolonized接触rsc突变体后,尽管其他动物成为后殖民延迟几个小时(Visick和Skoufos 2001)。这些结果表明,突变体被屏蔽殖民处于初级阶段,但是,他们可以偶尔旁路这个块,最终实现似乎是正常的殖民。在文化、rsc突变体没有表现出明显的缺陷在增长,能动性,或生物荧光诱导的时间和水平,为殖民特征被认为是重要的(Visick和Skoufos 2001)。因此,到目前为止没有线索rsc函数获得了表型观察到的文化。
rsc的顺序显示,它编码一种混合传感器类似ArcB激酶和bvg(图2 b) (Visick和Skoufos 2001)。这些蛋白质包含,除了守恒的残渣,作为网站的自身磷酸化,两个额外的域守恒的残留物(Asp)和他的预测是顺序和磷酸化,网站可以作为额外的监管(图2 b)。收到一个殖民信号,rsc预计autophosphorylate和磷酸转移至今未得到确诊的反应调节器,称为RscR,调节基因或活动必不可少的共生关系。
作为殖民信号,以及被rsc吗?很明显,存在许多可能性,包括除了细菌产生信号分子和海水,LO粘液的组件,和营养细胞表面信号。确定rsc的部分负责检测殖民共生信号交换信号将推进我们的理解。在许多情况下,传感器激酶蛋白的氨基末端周质部分接收环境信号(股票et al . 2000年)。例如,PhoQ沙门氏菌检测Mg2 +环境中通过其周质的领域;Mg2 +绑定到该域的结果的构象变化和失活反应调节器PhoP (Vescovi et al . 1997年)。rsc预计拥有~ 200周质的域残留物(Visick和Skoufos 2001);这个地区的大尺寸显示它可能扮演一个角色在rsc函数,可能信号检测。
除了潜在的周质的信号域,rsc包含一个输入域,称为不是。在其他蛋白质,不是检测信号等小配体,或光水平变化,氧浓度或氧化还原电位(泰勒和竹林1999)。是否不是域对信号检测的rsc殖民期间仍然是未知的。然而,从海水过渡到营养丰富的LO可能影响的能源地位诉fischeri细胞从而改变其氧化还原电位或氧浓度,可以感觉到的不是域。因此,调查rsc的PAS和周质的领域将富有成效的探索bacteria-host交互。也许每个域检测到一个不同的条件,允许rsc集成多个信号从鱿鱼环境调节殖民的开始。
同源响应监管机构的身份,RscR,和基因或蛋白质由rsc / R监管体系控制吗?在许多情况下,传感器激酶基因及其同源染色体响应监管机构联系,在某些情况下,基因控制的监管机构也在附近。这不是rsc的理由和基因编码的响应。诉的出现fischeri基因组测序项目(http://ergo。integratedgenomics.com/Genomes/VFI),使得它可以使用生物信息学RscR。使用序列已知的监管机构,我们有搜索和识别约40响应监管机构(Hussa Visick, unpubl。数据)。至少14出现分离的传感器激酶基因,从而代表RscR的最佳候选人。目前的工作旨在诱变处理这些候选人,问是否有突变体展览rscS-like殖民的缺陷。如果rscR编码DNA结合蛋白,那么新可用的DNA微阵列将用于探索rsc / R控制的调节子。识别目标的rsc / R监管也可能表明一个角色在共生启动调节子。一旦这些监管机构的目标(s)识别,实验旨在识别殖民信号可以制定。
gacA。在许多致病菌,双组分系统gac /毒性和主机协会调节表达特征,如假单胞菌的胞外酶的生产。(Heeb和哈斯2001)和能动性在沙门氏菌(2001年古蒂和ahm)。诉fischeri GacA主持人协会中也扮演了重要的角色。gacA表现出严重缺陷的突变体缺陷开始殖民:只有10%的动物成为殖民地,这些动物成为殖民展览减少近100倍的殖民(即。的细菌驻留在LO)(2003年惠斯勒和Ruby)。GacA的作用可能非常复杂。在文化,它与许多表型被认为是重要的共生关系,包括运动、营养收购,含铁细胞活动和发光(2003年惠斯勒和Ruby)。全球控制不同的特征,所有这些都有助于host-association,突出这样的监管机构的重要性共生进化的关联。与rsc / R,无论是信号还是基因/蛋白质GacA目标/ S是已知的。识别目标GacA监管,可能通过DNA微阵列实验,将有助于阐明这种调节器的作用对宿主共生和潜在揭示未知特征重要交互。
flrA。FlrA,转录监管机构有限序列相似性响应监管机构,主调节器的功能鞭毛生物合成(2003年米利根和Ruby)。鉴于共生起始运动性的绝对要求,要求FlrA似乎简单的突变导致缺乏鞭毛。然而,补充flrA突变体显示恢复运动性但不正常殖民:启动延迟和殖民的水平在48 h post-inoculation降低了10倍。
一种解释上述结果是鞭毛生物合成的时间和水平是最优的关键起始和殖民和这些特征并不是在补充压力恢复正常。支持这一假说,hyper-motile (hyper-flagellated)诉fischeri突变体也表现出严重的延迟启动殖民和缺陷水平的殖民24小时post-inoculation(密立根2002年和Ruby)。或者,一个同样可信的解释是,FlrA控制基因以外的其他参与鞭毛生物合成(密立根2003年和Ruby)也需要殖民。
几个non-flagellar基因似乎受FlrA(密立根2003年和Ruby)。一个基因被FlrA压抑,hvnC,编码一种蛋白质相关HvnA HvnB,两个分泌NAD + glycohydrolases诉fischeri中找到。然而,无论是hvnA还是hvnB出现所需殖民(Stabb et al . 2001);因此,FlrA-mediated hvnC监管的相关性尚不明确。第二个假定的霍乱弧菌kefB FlrA-repressed基因同源。在E。杆菌、KefB钾排出蛋白是重要的保护细胞免受有毒代谢物在生长在一个贫穷的碳源(弗格森et al . 2000年)。可能,诉fischeri KefB同族体从LO-specific毒素提供保护。
FlrA-repressed基因相关共生殖民?FlrA-controlled鞭毛,所需的启动,成为可有可无的殖民地的细菌。因此,一个开关在鞭毛基因转录与感应协调或通过FlrA non-flagellar基因的镇压。FlrA本身的监管可能在转录水平,类似于cAMP-CRP介导的控制权掌握在大肠杆菌鞭毛监管者flhDC (Soutourina et al . 1999年)。此外,有限的相似性FlrA响应监管机构表明其活动可以调节传感器通过磷酸化的激酶。未来的工作可能会专注于确定FlrA本身是转录控制,是否过度期间FlrA殖民影响转录水平或定时的假定的FlrA-controlled基因,这种基因本身是否促进(或干扰)殖民。
继续阅读:引用Mjt
这篇文章有用吗?