埃克曼抓图
1 o厕所
iooo
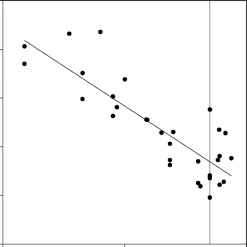
图1的密度底栖动物筛筛孔尺寸的函数在一系列湖泊从北方温带(r2 = 0.72, p < 0.000001)。垂直灰色线是500毫米网格通常用于大型底栖生物。
1 100 1000 10000浮游植物生产(g DM m 2一年级)
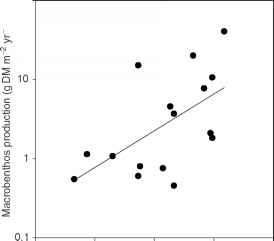
图3生产大型底栖生物和浮游植物产量之间的关系在一系列的湖泊(r2 = 0.34, p = 0.02)。改编自Kajak et al .(1980)的数据。
1 100 1000 10000净浮游植物生产量(g DM m 2一年级)
图3生产大型底栖生物和浮游植物产量之间的关系在一系列的湖泊(r2 = 0.34, p = 0.02)。改编自Kajak et al .(1980)的数据。
4 o i2o
有机沉积(g m-2yr2)
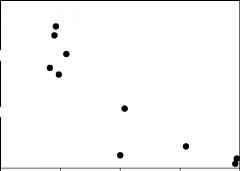
图2的比例zoobenthic生物量属于较小型底栖生物,作为一个功能的有机沉积在不同的网站深水层Paijanne湖、芬兰(r2 = 0.79, p = 0.0006)。改编自Hakenkamp et al。(2002),经过数据的Sarkka (1995)。
4 o i2o
有机沉积(g m-2yr2)
图2的百分比zoobenthic生物质属于较小型底栖生物、有机沉积的函数在不同网站的深水层Paijanne湖,芬兰(r2 = 0.79, p = 0.0006)。改编自Hakenkamp et al。(2002),经过数据的Sarkka (1995)。
的几个生态湖泊底栖动物的重要物种,一旦有地理分布范围小,特别是Dreissena spp。(斑马贻贝、双壳纲)、糠虾幼体(负鼠虾、甲壳纲动物),和几个小龙虾的物种。
因为湖泊底栖动物是如此的不同,很难概括其成员的生物特征。最大的动物在社区(双壳类和十足类,> 101克干质量)是更多
门 |
估计 |
典型的密度 |
数量 |
湖泊(没有。米2) |
|
的物种 |
||
在镜子湖 |
||
海绵动物门(海绵) |
4 |
NA |
刺细胞动物(一再出现的问题 |
2 |
100 - 1000 |
和水母) |
||
扁形动物门 |
40 |
1000 - 50000 |
(扁虫) |
||
纽形动物门 |
1 |
< 100 |
(海带) |
||
腹毛动物门 |
30. |
100000 - 1 000000 |
轮虫纲 |
210年 |
10000 - 250000 |
线虫纲 |
35 |
100000 - 1 000000 |
(蛔虫) |
||
环节动物门(蚯蚓, |
30. |
5000 - 50 000 |
水蛭) |
||
软体动物类(蜗牛、蛤) |
6 |
100 - 1000 |
Ectoprocta(苔藓 |
2 |
< 100 |
animicules) |
||
甲壳纲动物(的水蚤, |
70年 |
20000 - 200000 |
种子虾, |
||
桡足类, |
||
亲戚) |
||
螯肢动物门(螨) |
50 |
1000 - 10000 |
缓步类(water-bears) |
5 |
1000 - 50000 |
Uniramia(昆虫) |
120年 |
1000 - 50000 |
世界各地的许多湖泊包含相同的门和相当数量的物种作为镜子湖,但还没有充分的研究。
世界各地的许多湖泊包含相同的门和相当数量的物种作为镜子湖,但还没有充分的研究。

图4湖泊底栖动物的一些重要成员。成年人的身体长度给出了括号。(一)线虫(2毫米);(b) gastrotrich(0.2毫米);(c)蛭形轮虫(0.5毫米);(d) ploimate轮虫(0.1毫米);(e) tubificid寡毛纲动物(50毫米);(f) hydrobiid蜗牛(3毫米);(g) bivalveDreissena(20毫米);(h) unionid双壳类(75毫米); (i) acladoceran (1 mm); (j) an amphipod (10 mm); (k) a mysid shrimp (20 mm); (l) an ephemerid mayfly (20 mm); (m) a chironomid (10 mm); (n) the phantom midge Chaoborus (10 mm).
图4湖泊底栖动物的一些重要成员。成年人的身体长度给出了括号。(一)线虫(2毫米);(b) gastrotrich(0.2毫米);(c)蛭形轮虫(0.5毫米);(d) ploimate轮虫(0.1毫米);(e) tubificid寡毛纲动物(50毫米);(f) hydrobiid蜗牛(3毫米);(g) bivalveDreissena(20毫米);(h) unionid双壳类(75毫米); (i) acladoceran (1 mm); (j) an amphipod (10 mm); (k) a mysid shrimp (20 mm); (l) an ephemerid mayfly (20 mm); (m) a chironomid (10 mm); (n) the phantom midge Chaoborus (10 mm).
超过100亿倍的最小(轮虫和gastrotrichs ~ 10 ~ 9 g DM),所以这个社区跨越一个巨大的身体大小的范围。zoobenthic动物的寿命范围从不到一个星期(轮虫,gastrotrichs)几十年(双壳类)。一些物种已经艰难的长期休息阶段(海绵小芽,ectoproct休眠芽,水蚤类的ephip-pia)允许人口重建自己经过长期不利的时期,并协助被动传播之间的湖泊。各种物种探查沉积物(双壳类,tubificid寡毛纲动物,Chaoborus),滑动在湿地中(gastrotrichs, ploimate轮虫、扁形虫),附加(固着轮虫)或者我(一些chironomids属Cricotopus)水生植物,或爬或附着在固体如石头(Dreissena,许多腹足类)。
底栖动物包括物种suspension-feed浮游植物(许多双壳类和一些chironomids)或间质细菌(Pisidium),吃草底栖藻类(腹足类和许多轮虫),deposit-feed陈腐的碎屑和细菌(tubificid寡毛纲动物),在其他食肉动物底栖动物(odonates tanypodine chiro-nomids dicranophorid轮虫)或浮游动物(Chaoborus),吃树叶(小龙虾,chrysomelid甲虫)或根(dorylaimid线虫幼虫的甲虫Donacia)的植物,或分解的叶子落入水中的陆生植物(等足类动物,毛翅蝇)。不足为奇的是,被动悬架,喂食器如hydropsychid毛翅蝇和黑色的苍蝇在湖泊的发生率远比在流动的水域,大概是因为电流在湖泊太慢或不可靠的为他们提供食物。
底栖动物有几个适应处理低氧浓度发生在许多底栖环境。一些穴居动物(chironomids和蜉蝣)产生的电流将含氧水带入否则缺氧沉积物的洞穴。在几个分类群的物种(如水蚤类的,chironomids,腹足类)生产血红蛋白,使氧气在缺氧条件下运输。几个种类的线虫,蛭形轮虫,gastrotrichs, tubificid寡毛纲动物、桡足类、介形亚纲动物,chi-ronomid chaoborid浑身缺氧深海底沉积物的常见生产湖泊。这些动物显然生存延长缺氧通过使用厌氧代谢的物质如糖原或进入延长滞育。最后,许多的底栖动物(例如,许多昆虫和有肺的蜗牛)完全避免低溶解氧的问题,从空气中获得氧气。
方法的研究
湖泊底栖动物的一些成员是足够大的直接观察原位(例如,通过潜水或面具和通气管),但大多数物种必须收集和带进实验室学习。科学家们已经发明了一种广泛的装备收集底栖动物(图5)。多数情况下,科学家们用,去心器,扫网,关闭袋子或箱子,或陷阱收集动物或沉积物。通常,动物需要分开的沉积物或植物。这通常是通过通过筛洗样品,通常0.1 1毫米的网,有时跟着染色染料的样品如玫瑰红或检查样品在低倍显微镜下找到动物的帮助。或多或少的实验方法等密度梯度离心分离使用硅溶胶(例如,硅溶胶®)或应用程序的冰或温和热有时使用。
在湖泊底栖动物变异
底栖动物的主要特征之一是它的极端补缀的性质。底栖动物的丰度和物种组成变化在湖泊,在尺度上从厘米(复制样品在一个站点)公里,以及湖泊之一。众所周知,许多因素影响底栖动物的丰度和物种组成。
变化在湖泊
底栖动物的组成几乎总是与湖内水深变化很大。几乎每一个物种的丰富的底栖动物随水深(图6),和物种丰富度往往低得多的深水湖比接近岸边(图7)。许多研究也报道总数的变化或底栖动物生物量与水深在单独的湖泊。没有单一的模式的变化丰富或生物量与水深适用于所有湖泊,因为许多机制,链接水深zoobenthic社区不同强度在湖泊和产生一个广泛的模式在不同的湖泊。
最重要的因素导致zoobenthic成分和数量随水深包括溶解氧、有机质输入的数量和质量,温度,沉积物粒度和压实,干扰,与深度有关的生物相互作用。所有这些因素的相对贡献及其交互在推动zoobenthic尚未进行社区结构。在分层的湖泊通过分层,hypolimnetic溶解氧的浓度下降,并且经常在沉积物表面达到零结束的夏天。只有少数几个物种的动物能容忍缺氧(< 2 mgl_1)或缺氧条件下对于任何一段时间。常常伴随着缺氧也是有毒的硫化氢对大多数动物。因此,溶氧不足的沉积物表面可能是一个主要原因的垂直zona-tion深海底沉积物的底栖动物和较低的物种丰富度。然而,下降丰富经常发生远高于氧消耗的深度(图7),所以氧气以外的因素必须是重要的。尽管一些zoobenthic物种可以容忍缺氧,这些物种可能是丰富的,所以低溶解氧不一定降低zoobenthic密度和生物量。即使在无层理的湖泊,温暖时期,无风的天气可能会减少混合足以导致短期消耗氧气的沉积物表面和底栖动物的灾难性损失。这样短暂的分层和氧气耗尽死亡几乎整个人口^ 2500吨(干质量)的蜉蝣Hexagenia浅盆地西部的1953年夏末伊利湖。
温度分层的深水层比epilim-nion湖要低得多,更稳定。这些低温缓慢的底栖动物的代谢率,可能无法满足阈值许多物种的生长和繁殖,并从深海底沉积物可能排除许多物种。
底栖动物的食物基地距海岸线变化,深水层。水生植物和附着藻类增长只有在较浅或清水。在深水中,新鲜phyto-plankton补充剂老沉砂洗的分水岭滨海区,有机质的数量和质量的深海底沉积物深湖随着水深可能下降。沉没的沉积有机质在湖床高度不均匀,
样品室
氯丁橡胶
氯丁橡胶
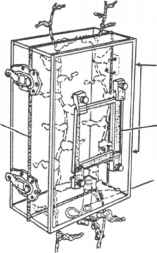
铰链
采样端口
出口管
铰链
采样端口
出口管
样品室
关闭触发
关闭触发
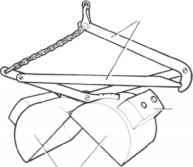
重量
大白鲨
重量
大白鲨
关闭触发
吸盘
气缸打开
关闭触发
吸盘
气缸打开
-可拆卸缸u
结束了
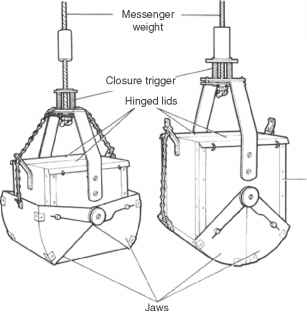
图5的四个取样器常用的定量研究湖泊底栖动物。(一)埃克曼抓住;(b)PONAR抓住;(c)唐宁箱取样器vegetation-dwelling无脊椎动物;(d) Kajak-Brinkhurst去心器。改编自唐宁(1984,1986)。
根据波能量和底部的斜率。似乎合理的假设的数量和质量的食物通常是最高的在沿岸带和沉积地区,并导致zoobenthic生物量的变化(图8)。
沉积物粒度和异质性也随水深,通常从一个高度的马赛克的粗粒沉积物在浅水区的单调的平原在深水层细粒度沉积物。因为许多机制,底栖动物与他们的沉积物,沉积物的变化必须对底栖动物的种类有很大影响,可以住在不同的深度在一个湖泊。高异质性浅水沉积物可能是一个主要的原因在沿海底栖动物物种丰富度高。
干扰从波浪冲刷、ice-push或水位波动常常引起显著的区域

Pallasea quadrispinosa
股!区
比提尼亚触手纲
Valvata piscinalis
比提尼亚触手纲
Acellus aquaticus
Dreissena polymorpha
Pallasea quadrispinosa
Valvata piscinalis
Acellus aquaticus
Dreissena polymorpha

沿海大型植物区
1阿 |
1 1 1 1 1 1 |
1 |
||
Theodoxus丁 |
一个 |
T 2 S 1 O |
||
1阿 |
} |
|||
1阿 |
Gammarus lacustris |
我 |
T 2 S 1 O |
|
是 |
1 |
|||
是 |
Oulimnius穴居人^ |
1阿 |
||
1阿 |
Nemoura avicularis Tinodes waeneri |
我 |
100年50 |
|
三世 |
||||
1阿 |
Sphaerium角质层 |
* |
是 |
|
是 |
Hydroptila femoralis |
- - - - - - |
是 |
|
Centroptilum |
T ^ |
|||
luteolum |
是 |
|||
J - o |
Gyraulus冠 |
» |
1阿 |
|
Eurycercus |
+ |
Ts |
||
lamellatus |
J - o |
|||
Ts |
||||
1阿 |
Micronecta poweri poweri |
1 |
1阿 |
|
滨海区
股!区
亚沿岸带
沿海大型植物区
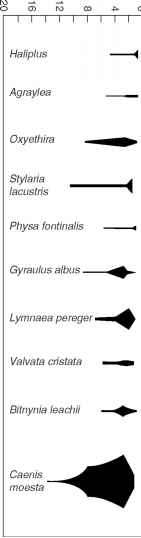
图6底栖动物的分布和水的深度富营养的湖Esrom,丹麦。改编自约(2004)。
沿岸带w
5”
3”
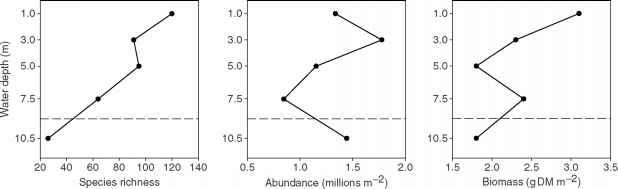
图7中物种丰富度、丰度和生物量的底栖动物(包括所有后生动物)水深的函数在镜子湖,一个小湖在新罕布什尔州徒劳无益。冲水平线显示氧气在沉积物表面的深度低于1毫克l1夏末。改编自不走正路的人(1985)。
图7中物种丰富度、丰度和生物量的底栖动物(包括所有后生动物)水深的函数在镜子湖,一个小湖在新罕布什尔州徒劳无益。冲水平线显示氧气在沉积物表面的深度低于1毫克l1夏末。改编自不走正路的人(1985)。
Epilimnetic沉积
Epilimnetic nondepositional
Hypolimnetic沉积
Hypolimnetic nondepositional
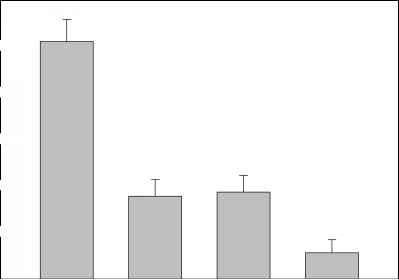
图8的意思(±1)生物质(湿质量)macrobenthic动物在不同网站Memphremagog湖,魁北克,作为温度的函数和沉积机制。改编自拉斯穆森和罗文(1997)。
Epilimnetic沉积
Epilimnetic nondepositional
Hypolimnetic沉积
Hypolimnetic nondepositional
图8的意思(±1)生物质(湿质量)macrobenthic动物在不同网站Memphremagog湖,魁北克,作为温度的函数和沉积机制。改编自拉斯穆森和罗文(1997)。
减少zoobenthic密度和多样性靠近水边,尤其是在大湖泊和水库。
最后,许多生物的相互作用影响底栖动物与深度有关的。鱼捕食数量可以有非常大的影响,大小,和底栖动物的物种组成(图9)。鱼的种类和数量的变化从沿海深水层;事实上,缺氧深海底沉积物可能是免费的从鱼捕食。同样,密度和类型的植物,这很重要,为附件提供表面,躲避捕食,底栖动物的食物,与水深变化明显,可以推动大型底栖动物的变化。因此,生物相互作用构成的湖中的底栖动物的变化。
最底栖动物附近发现沉积物表面(图10),大概是因为食物(沉没ben-thic藻类和浮游植物)和氧气是最可用的。然而,在湖泊沉积物动物可以找到更深,有时达到深度超过50厘米。Tubificid oligo-chaetes进料口的沉积物,因此很容易理解为什么他们达到深层湿地接口,但其他底栖动物生活的活动深湖中的沉积物(例如,一些candonid介形亚纲动物,虫和蛭形轮虫)不太理解。

10 15 20 25
图9的依赖macrobenthic生物质(湿质量)在芬兰Pohjalampi湖鱼捕食。每个点是一年在鱼类种群的长期实验操作。改编自Leppa et al。(2003)。
10 15 20 25
图9的依赖macrobenthic生物质(湿质量)在芬兰Pohjalampi湖鱼捕食。每个点是一年在鱼类种群的长期实验操作。改编自Leppa et al。(2003)。
表面密度的密度(%)50 100 150 200
表面密度的密度(%)50 100 150 200
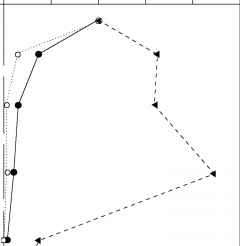
图10底栖动物的垂直分布在镜面湖的沉积物,新罕布什尔州。三行显示所有的底栖动物的分布,一个surface-dwelling分类单元(monogonont轮虫),和一个分类单元,生活更深的沉积物中(蛭形轮虫Rotaria tridens)。修改从流浪者(1985)。
变化在湖泊
有机质输入,尤其是浮游植物生产、影响底栖生物的种类和数量动物在湖泊(图11)。通常,湖泊与高有机质输入支持更多的底栖动物(图3和11),尽管有证据表明,高投入的有机物会减少数量的底栖动物,可能通过增加缺氧沉积物的面积。底栖动物的种类变化与湖泊生产力。事实上,最早的系统分类湖泊的生产力是基于深底底栖动物的组成。
湖的形态测量学对zoo-benthic社区有很强的影响。小,浅水湖泊倾向于支持更高的底栖动物的密度比
2500年
2000 -
1500 -
500 -
0.001
0 20 40 60 80 100 120
的背影,浮游植物生物量(^ gl-1)
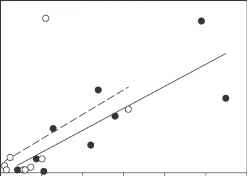
图11显示例子的依赖浮游植物的底栖动物。片脚类动物的上面板显示数字Monoporeia亲近种Erken湖,瑞典,的函数中重要的浮游硅藻Aulacoseira前一年(r2 = 0.54, p = 0.0008)。较低的面板显示的生物质chironomids 4在巴拉顿湖的两个网站,匈牙利,浮游植物生物量的函数在去年夏天(黑色圆圈,r2 = 0.66, p = 0.008;白色圆圈,r2 = 0.20, p = 0.23)。改编自约翰逊和Wiederholm(1992)和Specziar: (2001)。
2000 -
1500 -
500 -
-
")
- 0.01 - 0.1 Aulacoseira biovolume (mm3l-1)
")
0 20 40 60 80 100 120
的背影,浮游植物生物量(^ gl-1)
图11显示例子的依赖浮游植物的底栖动物。片脚类动物的上面板显示数字Monoporeia亲近种Erken湖,瑞典,的函数中重要的浮游硅藻Aulacoseira前一年(r2 = 0.54, p = 0.0008)。较低的面板显示的生物质chironomids 4在巴拉顿湖的两个网站,匈牙利,浮游植物生物量的函数在去年夏天(黑色圆圈,r2 = 0.66, p = 0.008;白色圆圈,r2 = 0.20, p = 0.23)。改编自约翰逊和Wiederholm(1992)和Specziar: (2001)。
大,深的湖泊。这种模式可能至少有三个原因:(1)植物,支持密集多样的zoobenthic社区,在小最丰富,浅水湖泊;(2)浅水湖泊不太可能比深湖泊分层,导致更高的溶解氧浓度在沉积物表面;(3)沉没浮游植物(和其他食物残渣)不太可能被降级之前在浅水湖泊沉积物表面比在深湖。
温度影响湖泊底栖动物的种类和数量。热带湖泊支持不同的物种(或比温带湖泊北极湖泊等),尽管集团chironomids, Chao-borus,寡毛纲动物丰富广泛纬度的范围。温暖的湖泊倾向于支持zoobenthic密度高于做冷湖。
水化学几个方面有很强的对底栖动物的影响。盐水湖含有独特的社区的底栖动物。非常咸湖可能包含几个种底栖动物,尽管总体丰富或底栖动物生物量可能不会减少。软水湖泊的低pH值和低钙通常包含不同的物种比硬质水湖泊,并且经常很穷在甲壳类动物(软体动物,介形亚纲动物),需要补充钙来构建他们的贝壳。因此,最近的酸化大气沉积有重要的对底栖动物的影响。高浓度的溶解有机物在湖水可以减少大量的底栖动物。
最后,差异在湖泊生物相互作用有很强的对底栖动物的数量和种类的影响。只有少数是众所周知的例子。Fishless湖泊通常包含大、活跃的无脊椎动物(例如,游泳或爬行昆虫),迅速消除如果鱼了。小龙虾和大双壳类可以在其他底栖动物有很强的效果,已被证明当入侵湖泊的外星小龙虾和Dreis-sena造成非常大的底栖动物群落的变化。似乎其他自然的生物相互作用是负责整个湖泊底栖动物的变化。
湖泊底栖动物的角色
二次生产的底栖动物大多数湖泊适度的(表2),能量流在底栖动物肯定小得多,通过微生物群落。然而,底栖动物扮演重要角色湖泊生态系统,甚至在人类事务。一般来说,底栖动物的生态角色可以定义为食物链或nontrophic角色。
参与者在湖食物网,底栖动物消耗浮游植物,水生植物,动物,细菌和碎屑。消费比例可高到足以控制这些食物资源的数量和成分。最著名的例子包括底栖动物的食用浮游生物。Suspension-feeders如Dreis-sena、海绵和水蚤类动物可能丰富足以降低浮游植物生物量,或改变其组成(图12)。同样,底栖动物,吃浮游动物(Chaoborus糖虾)可以对浮游动物群落产生强有力的控制,
湖 |
平均深度(米) |
净有机输入 |
的生产 |
的生产 |
(g C m2yr1) |
底栖动物(%) |
浮游动物(%) |
||
苔原池塘,正义与发展党 |
< 1 |
26 |
7 |
0.8 |
马里昂,公元前 |
2 |
110年 |
3 |
0.9 |
Myvatn、冰岛 |
2 |
330年 |
6 |
1.3 |
Wingra, WI |
3 |
610年 |
0.4 |
4 |
乌克兰基辅水库, |
4 |
280年 |
6 |
7 |
Rybinsk水库、俄罗斯 |
6 |
93年 |
0.3 |
2 |
镜子,NH |
6 |
49 |
12 |
5 |
红色,俄罗斯 |
7 |
140年 |
1 |
7 |
Findley佤邦 |
8 |
12 |
6 |
2 |
Naroch、白俄罗斯 |
9 |
160年 |
0.8 |
5 |
Mikotajskie、波兰 |
11 |
260年 |
2 |
21 |
Esrom、丹麦 |
12 |
160年 |
6 |
7 |
Paajarvi、芬兰 |
14 |
60 |
3 |
12 |
Dalnee、俄罗斯 |
32 |
260年 |
1 |
22 |
最底栖生物数据排除较小型底栖生物,因此低估了。由于方法论上的差异研究,数据只是大约可比。编译,还可以从许多别的来源。
最底栖生物数据排除较小型底栖生物,因此低估了。由于方法论上的差异研究,数据只是大约可比。编译,还可以从许多别的来源。
分枝的影响通过生态系统(图16)。尽管相对较少的底栖动物(小龙虾、plant-parasitic线虫和一些水生昆虫)吃或破坏水生植物,它们能很强烈地影响大型植物生物量和物种组成;的确,底栖动物被用于生物防治公害杂草如耆草属植物。不太了解在附着藻类、底栖动物的影响细菌,和碎屑,尽管这些是最底栖动物的主要食物。此外,一些底栖动物高度专业化的饮食,因此可能对食物资源有选择性的影响。底栖动物可能经常控制附加在湖泊藻类的数量和类型,以及似乎大多数的碎屑颗粒通过至少一个底栖动物在被埋在湖底。因此,
没有贻贝
没有贻贝
-

- 贻贝补充道


绿藻NXX硅藻
蓝藻其他
图12浮游植物的数量和组成的变化在两个小荷兰湖泊实验后添加Dreissena polymorpha湖泊之一。每个圆的面积成正比的biovolume每个湖浮游植物。改编自里德的数据等等。(1993)。
有充分的证据表明,底栖动物可以控制食物资源的数量和特征在湖泊,尽管我们还不知道多久,在什么条件下发生如此强烈的控制。
底栖动物时释放无机氮、磷等营养物质消耗的食物。这样的营养再生可能是重要的浮游植物和附加固着生物nutrient-limited湖泊。
zoobenthic生产主要的命运是被吃掉,是否由其他成员的底栖动物,鱼,或陆地掠食者(表3)。因此,底栖动物作为一个重要的链接到高营养水平。几乎所有木架房屋鱼在某种程度上取决于底栖动物和底栖动物的主要食物是许多鱼类(图13)。大量的鸟类、蝙蝠、蜘蛛和其他食肉动物可能会吸引到湖泊或湖滨以底栖动物为食,包括新兴昆虫。
虽然已经习惯把浮游生物和底栖生物属于单独的食物网,许多物种的底栖动物直接或间接依赖浮游生物为食,也有很强的浮游生物和底栖动物之间的互惠链接。的远洋和底栖生物区湖是联系在一起的许多强大的连接(图14),作为一个集成系统和功能。
底栖动物是沉积物的主要nontrophic角色混合(生物扰动作用)。三个活动重要:(1)喂养,尤其是通过传送带喂食器如tubificid寡毛纲动物,饲料在深厚层沉积物表面的沉积物和把粪球;(2)洞穴建筑和通风,通过蜉蝣蜉蝣和一些chironomids;(3)普通运动活跃的大动物,如Chaoborus和unionid双壳类。生物扰动作用混合沉积物和增加
湖 |
生产(gDM m 2 yr1) |
命运的生产(%) |
|||
无脊椎动物捕食 |
鱼捕食 |
鸟类捕食 |
出现 |
||
Myvatn、冰岛 |
42 |
5-52a |
43 |
9 |
48 |
镜子,NH |
14 |
80年 |
15 |
nd |
25 |
Paajarvi、芬兰 |
3.8 |
50 |
40 |
nd |
6 |
Batorin、白俄罗斯 |
2.6 b |
46 |
194年 |
nd |
nd |
Naroch、白俄罗斯 |
2.6 b |
22 |
42 |
nd |
nd |
Ovre Heimdalsvatn、挪威 |
2.4摄氏度 |
25-28a |
70年 |
2 |
3 |
Myastro、白俄罗斯 |
0.8 b |
72年 |
272年 |
nd |
nd |
数据是近似的,所以百分比不总和为100%;nd = aEstimated差异。生长季节;大型底栖生物。cMacrobenthos。
不确定。数据从各种来源。
数据是近似的,所以百分比不总和为100%;nd = aEstimated差异。生长季节;大型底栖生物。cMacrobenthos。
不确定。数据从各种来源。
T3 O
-
.")
- 图13的底栖动物的饮食常见物种北美东北部冷漠的湖泊。“间接”的底栖动物是吃鱼的消费由底栖动物。改编自编制的数据Vadeboncoeur et al . (2002)。
.")
溶解物质的交换(如氧气、铵)在上覆水和沉积物孔隙水之间。这些活动可以有大影响营养再生和循环和埋葬的毒素。此外,建造甲壳的底栖动物(主要是大型双壳类)可以改变底栖生物栖息地的特征通过他们的生活积累和死贝壳。shell积累,这样可以达到质量> 1公斤m ~ 2,作为其他动物栖息地或产卵网站,以及改变湿地生物地球化学转换。
应用问题
一些最重要的人类和野生生物的疾病是由底栖动物。其中最主要的是疾病引起的吸虫(或吸虫),淡水螺,偶尔十足类,作为中间宿主。在一个典型的生命周期(图15),鸡蛋从终宿主孵化成幼虫游泳(miracidia),寻找并穿透一个水生蜗牛。许多种类的淡水螺作为影响人类的各种吸虫的宿主,牲畜,和野生动物。在蜗牛进行开发后,第二个(尾蚴)漂流的可以直接进入终宿主主机在水中时,进入第二中间宿主(鱼或十足类动物),或被包在囊内水生植物。吸虫从第二中间宿主或水生植物进入终宿主,当生鱼,十足类,或者食用水生植物。成人吸虫可能生活在肠道、肝脏、血管,或肺的人类和其他脊椎动物,而且往往引起慢性、退行性疾病。schisto-somiasis snail-borne疾病,最重要的使人衰弱的疾病引起的裂体吸虫属血吸虫,影响^ 2亿年人类在整个热带地区。其他重要snail-borne疾病包括肝吸虫(特别是肝片吸虫和Opisthorchis sinensis),大危害绵羊和牛,以及影响数以百万计的人,和各种肠道或肺吸虫,这影响到世界各地的许多人。吸虫类使用淡水蜗牛作为中间宿主也影响许多其他脊椎动物比人类。事实上,吸虫尾蚴的鸟类使用的主机有时探查人类。虽然这样cercariae不正常发育在人类身上,他们会引起皮肤刺激称为游泳的痒,从而限制娱乐湖泊的使用。acanthocephalans是另一个重要的寄生虫由淡水底栖动物。成年acanthocephalans许多鱼类和其他水生脊椎动物的肠道寄生虫,和底栖甲壳类动物通常作为中间宿主。
人为自然栖息地的变化有时会增加疾病的问题(例如,蓄水池增加了血吸虫病的流行),和生态干预措施(例如,栖息地管理、介绍捕食者或其他生物控制)可以实现疾病控制综合计划的一部分。
其他底栖生物物种是害虫。污染物种如Dreissena、蚬,偶尔海绵和ectoprocts阻水的摄入量,可能迫使核电站运营商使用机械清洗,bio-cidal化学物质,或管道涂料保持水流。看来大的事chironomid蚊虫可能的一个主要原因花粉热在世界的某些地方和哮喘。质量的事木架房屋昆虫(主要是蜉蝣蜉蝣或chironomid chaoborid浑身)可能是如此之大以至于导致交通危险,生产料堆需要清理的昆虫尸体重型设备,甚至短路nb雷竞技官网 。
尽管有一些兴趣biomanipulation增加的积极影响底栖动物,或减少其负面影响,这些努力并没有进行的远洋区现在,biomanipulation广泛实践。至少有三种类型的biomanipulation被认为涉及底栖动物。因为底栖生物
Suspensionfeeding底栖生物
Deposit-feeding底栖生物

Suspensionfeeding底栖生物
Deposit-feeding底栖生物
淹没的植被
淹没的植被
图14主要湖社区之间的相互作用(不走正路,2006)。箭头显示的虚拟方向控制;注意,许多交互箭头沉积物和远洋区之间的交叉。
一个= =诊断感染性阶段阶段
孢囊在蜗牛^ ^(一代又一代)
蜗牛,Cercariae发布成水和自由游动的o g
鸡蛋孵化释放miracidia
在渗透,成为schistosomulae Cercariae失去尾巴
问循环
一个= =诊断感染性阶段阶段
孢囊在蜗牛^ ^(一代又一代)
蜗牛,Cercariae发布成水和自由游动的o g
在渗透,成为schistosomulae Cercariae失去尾巴

问循环
迁移到门户在肝脏和血液成长为成年人问
成对成虫迁移到:问B肠系膜小静脉肠/直肠(产卵循环到肝脏和粪便)C膀胱静脉丛
图15生命周期的三种血吸虫血吸虫。来自美国疾病控制中心。
迁移到门户在肝脏和血液成长为成年人问
成对成虫迁移到:问B肠系膜小静脉肠/直肠(产卵循环到肝脏和粪便)C膀胱静脉丛
图15生命周期的三种血吸虫血吸虫。来自美国疾病控制中心。
suspension-feeders可能控制浮游植物生物量和组合,一些研究试图管理麻烦浮游植物通过增加底栖生物种群suspension-feeders(图12)。为鱼Ben-thic动物是有价值的食物,所以渔业管理者往往推出了大型底栖动物
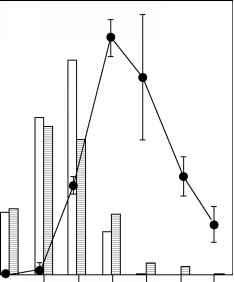
(小龙虾和负鼠虾糠虾幼体)湖泊鱼类种群的增长率或生物量增加。支持了这样的介绍很少仔细分析可能的影响,并介绍饲料无脊椎动物有时会导致不良的和不可预见的影响(图16)。最后,
1987年
1989年
图16不良影响之后的引入负鼠虾糠虾容易受骗的湖,蒙大拿。红大麻花鱼鲑鱼和鹰峰计数数据从支流流使用的产卵的鲑鱼。改编自斯宾塞et al。(1991)。
1979 1981 1983 1985
一年
1987年
1989年
图16不良影响之后的引入负鼠虾糠虾容易受骗的湖,蒙大拿。红大麻花鱼鲑鱼和鹰峰计数数据从支流流使用的产卵的鲑鱼。改编自斯宾塞et al。(1991)。
有一些试图提高底栖动物种群的控制人口的动物传播疾病。也许,最突出的例子是捕食者或竞争对手的增加携带schistosomia-sis的蜗牛。显然,biomanipulation湖泊利用底栖动物仍然处于起步阶段。
和其他淡水植物和动物一样,人类活动伤害了一些湖泊底栖动物的物种,它们现在已经灭绝或濒危。几个活动尤为有害。大型水改道造成一些湖泊成为过度咸甚至完全枯竭,危及或消除底栖动物,以前住在那里。许多湖泊已被毒素污染严重的工业和其他来源或沉积物从贫穷的土地利用方式。营养负荷的化肥或国内浪费增加了许多湖泊的生产力,导致缺氧沉积物,明显影响底栖动物。许多外来物种(运动鱼类、水生植物和无脊椎动物)被引入世界各地的湖泊有强烈的对底栖动物的影响。会计的总结这些有害的活动对底栖动物的影响不存在,但许多种群个体湖泊已濒危或取消。在古代支持特有种的湖泊底栖动物,当地消灭将转化为全球物种灭绝。古代湖泊特有鱼类的灭绝是众所周知的,大概,这样灭绝发生在底栖动物。
几个种底栖动物从湖泊或水库供人类使用。重要的野生渔业或各种淡水养殖种群十足类(小龙虾、对虾、蟹)分布湖泊、池塘,世界各地的湿地。几个大型双壳类的几千年来一直收获的食物、珍珠、珍珠母,虽然这些动物是最大的渔业在湖泊河流而不是。特别是在十九世纪,大量(> 1000万只/年)药用水蛭(水蛭从野生medici-nalis)拍摄。湖泊底栖动物的其他成员(成人chaoborid蚊虫,鸡蛋和成人corixid bug,蜗牛)收获大量对人类在本地食物。
术语表
海底生物的社区生活在表面(例如,沉积物,植物)在水生生态系统。
生物扰动作用,沉积物混合生物的活动引起的。
大型底栖生物的底栖动物大到足以被保留在粗筛(通常是~ 0.5 mm网)。
较小型水底生物-底栖动物太小被保留在粗(通常~ 0.5 mm网)
筛,但足以被保留在一个晴朗的(通常^ 0.05毫米网)筛。
底栖动物,动物的社区生活在表面(例如,沉积物,植物)在水生生态系统。
参见:Biomanipulation水生生态系统;
沿岸带;营养动力学在水生生态系统。
进一步的阅读
Brinkhurst RO(1974)湖泊的海底。纽约:圣马丁出版社。
唐宁JA(1984)抽样运行水域的海底。:喝JA和Rigler FH (eds)。手动方法评估次级生产力的新鲜水,第二版。,87 - 130页。牛津大学:布莱克威尔。
唐宁J(1986)回归估计技术的附生植物的无脊椎动物的种群数量。淡水生物学16:161 - 173。
Hakenkamp CC,莫林,流浪者DL(2002)的功能淡水的重要性较小型底栖生物。:梯级SD,罗伯逊,Schmid-Araya JM (eds)。淡水较小型底栖生物:生物学和生态学,页321 - 335。Backhuys。
哈钦森通用电气(1993)论述湖沼学,第四卷,底栖动物。纽约:威利。
约翰逊RK Wiederholm T (1992) Pelagic-benthic耦合振荡——硅藻年际变化对人口的重要性Monoporeia亲近种。湖沼学和海洋学37:1596 - 1607。
约下午(2004年)底栖无脊椎动物。:奥沙利文PE和雷诺兹CS (eds)。湖泊手册,卷1。湖沼学和湖沼生态,页341 - 416。莫尔登:布莱克威尔。
Kajak Z, Bretschko G、Schiemer F和C (1980) Zoo-benthos桑德琳。:Lecren埃德和Lowe-McConnell RH (eds)。淡水生态系统的功能,页285 - 307。剑桥:剑桥大学出版社。
Leppa M, Hamalainen H, Karjalainen J(2003年)底栖无脊椎动物的反应,整个湖水biomanipulation。Hydrobio-logia 498: 97 - 105。
梅里特RW,康明斯千瓦,Berg MB (eds)(2007)介绍北美的水生昆虫,第四版。迪比克:肯德尔/打猎。
拉斯穆森JB Kalff J (1987)实证模型在湖泊zoobenthic生物量。加拿大渔业和水产科学》期刊上44:990 - 1001。
拉斯穆森JB和罗文DJ(1997)波速阈值对细颗粒泥沙堆积在湖泊、zoobenthic生物量及其影响和构成。《北美Benthological社会16:449 - 465。
里德H, bij de Vaate, Noordhuis R(1993)潜在的斑马贻贝(Dreissena polymorpha)对水的质量管理。:Nalepa TE和Schloesser DW (eds)。斑马贻贝:生物学、影响和控制,439 - 451页。佛罗里达州波卡拉顿的:刘易斯出版商。
梯级SD,罗伯逊,Schmid-Araya JM (eds)(2002)淡水较小型底栖生物:生物学和生态学。莱顿:Backhuys出版商。
Sarkka J(1995)深底较小型底栖生物在两个大湖:污染和水深的影响差异。档案毛皮Hydrobiologie 132: 453 - 493。
Specziar和用途:L(2001)长期动态的巴拉顿湖的chironomid动物和浮游植物生产的依赖。档案毛皮Hydrobiologie 152: 119 - 142。
斯宾塞CN、麦克勒兰德BR和斯坦福JA(1991)虾长袜,鲑鱼崩溃,和鹰位移。生物科学41:程度。
不走正路的D(1985)的底栖生物micrometazoans镜子湖,新罕布什尔州。档案毛皮Hydrobiologie Supplementband 72: 287 - 426。
不走正路的D(1991)视角大小湖泊底栖动物的结构,其原因,其后果。北美Benthological社会杂志》10:210 - 221。
不走正路的DL(2006)的底栖动物群落tidal-freshwater哈德逊河河口。JS: Levinton和Waldman小(eds)。哈得逊河河口,页266 - 278。剑桥大学出版社。
索普JH和Covich美联社(eds)(2001)北美淡水无脊椎动物生态学和分类,第二版。圣地亚哥:学术出版社。
Vadeboncoeur Y,范德Zanden MJ,洛奇DM(2002)把湖起来:将底栖生物通路湖食物网模型。生物科学52:44-54。
继续阅读:湖泊和水库的富营养化
这篇文章有用吗?
读者的问题
-
迪莉娅1年前
- 回复
-
J1年前
- 回复