滨海区
雷竞技raybet最新版下载J彼得斯和D米洛奇,巴黎圣母院,巴黎圣母院大学,美国
©2009爱思唯尔有限公司保留所有权利。
介绍
的滨海区湖是近岸的接口和陆地生态系统之间的更深远洋区的湖。是区域至少百分之一的光合活性光(400 - 700 nm)进入水到达沉淀物,使初级生产者(大型植物和藻类)蓬勃发展。滨海区在结构上和功能上的一个重要组成部分大多数湖泊有几个原因。首先,地球上大部分湖泊都很小,因此,滨海区包括一个大湖泊总面积的比例(图1)。第二,作为一个接口,滨海区影响物质的运动和处理从地面径流流入湖泊,地下水,或流连接,从而影响该区域的物理和生物过程的其余部分湖泊生态系统。第三,沿岸带通常是最具生产力的区域的湖泊,尤其是水生植物和无脊椎动物。最后,人类利用水生系统(游泳、钓鱼、划船、发电、灌溉、等等)往往集中在沿海地带。
在第一部分,沿岸带内的物理结构和营养动力学描述。在第二部分中,沿岸带的生物体之间的相互作用进行了讨论,除了滨海和陆地生态系统之间的相互作用,并在沿海和远洋区。在最后的部分,人为影响在滨海区。
影响因素的物理结构和养分动态滨海区
许多特征确定的百分比滨海区和湖类型的湖底基板。滨海区面积和基质类型,反过来,影响加工的营养物质,矿物质和有机质,因此整个湖的功能。
沿岸带的物理结构
分带,滨海区可分为上,中间,和较低的区域,从海岸线地区喷洒波滨海带的底部,低于光不能穿透
(图2)。紧急植被是根植于上层沿岸带;漂浮植物发现中间沿岸;和submergent植被通常生长在沿岸。littoriprofundal,居住着藻类和自养细菌,下面是一个过渡区较低的沿海区域。下面这个过渡区,微粒永久定居到深水层因为风或对流能源是不够的在这些深度resuspend粒子。沿岸带深度通常对应于夏季变温层深度分层的湖泊。
栖息地与沉积物的均匀分布在深水层,滨海带的栖息地和沉积物分布异构补丁(图3)。
沉积物中粒子的大小范围从非常好的有机和无机粒子(淤泥或淤泥)大鹅卵石和巨石。大型植物和倒下的树木通常提供垂直基板在沿岸带(指“参见”部分)。丰度和分布在沿岸带调和了丰富的栖息地,生物的多样性,和交互。例如,鹅卵石基质提供避难所从鱼捕食小龙虾;相比之下,精细有机基质支持大型植物的生长为无脊椎动物提供庇护,浮游动物和幼鱼。无脊椎动物丰度和组成不同在不同的衬底。整体底栖无脊椎动物多样性大于异构沿海地区较均匀的深底区域。稍后解释,栖息地在沿岸带的类型取决于湖的形态测量学周围的景观,风的模式和营养负荷,湖。
湖泊形态测量学的形态学特征影响栖息地在沿岸带的种类包括湖地区,深度,海岸线曲折度和水下斜坡。一个很大程度上决定了湖湖形态测量学的起源。例如,湖泊形成通过构造或火山活动通常是非常大的,陡峭的湖泊以最少的沿海地区,而冰川湖泊和水库经常有复杂盆地形状和大的沿海地区。
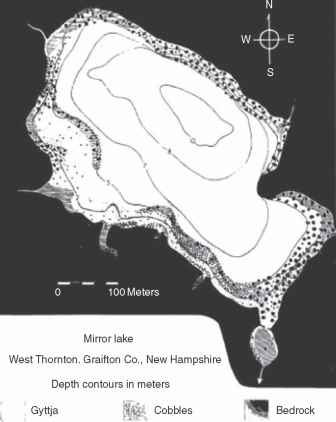
与大湖区湖泊深度比例,更多的蜿蜒的海岸线、更复杂的深度测量法,和浅倾斜的盆地将有一个更大的百分比沿岸带与远洋区。例如,浅水湖泊大表面积大滨海区因为光线能穿透到湖的很大一部分地区沉积物。
湖泊形态测量学特征也影响沿岸带内发现的基质类型。陡峭的倾斜的滨海地区通常有岩石/鹅卵石基质,和地区逐步斜率可以由细沉积物有或没有大型植物(图3)。海岸线曲折度高的湖泊有更多的海湾与大型植物生长在沙子或淤泥与圆形的湖泊相比,由于波浪作用是减少保护海湾、允许的积累好
“o rn
“哦,
沿海主要远洋主导远洋比滨海带的区域
图1的数量湖泊的世界由沿海和远洋区。修改从吉姆RG(2001)湖沼学:湖和河流生态系统。纽约:爱思唯尔,学术出版社。
有机沉积物,营养素和矿物质,建立大型植物。
周围的景观湖周围的地形、地质的土地影响水的运动,营养有关,矿物质和有机质在沿岸带。地表径流和地下水的相对贡献一个湖取决于水渗透和传播率周围的土壤,陆地植被的生产力,斜率和水系密度的分水岭。
海拔和水文流定义在景观湖的位置。高的景观,湖泊往往是小渗流湖泊,这是美联储主要由降水和地下水。大排水湖泊,美联储的地表水,地下水,沉淀,往往是低的格局(图4)。湖泊景观往往有较大的降低,更有效率的沿海地区,因为大分水岭输入的营养物质,矿物质,和溶解或微粒有机材料,从地表水和流连接。这种材料的输入增加缓冲能力降低酸化的影响(能力)和大型植物和无脊椎动物的丰度和多样性像蜗牛住在大型植物。同时,湖泊景观通常有一个更复杂的盆地低深度测量法,也可以提高沿海地区。
风基质在沿岸带风的函数如获取和接触。获取整个湖是风吹的距离。迎风和李的湖会有截然不同的底物的特征。风引起的波浪作用越强,越微粒将暂停,最终沉积的深水层湖泊沿岸带衬底越将岩石的特征。波浪作用也将减少蜿蜒的海岸线和大型植物如前所述。
-


- 图2滨海带的分带。列出相关波条件、底物和无脊椎动物。上游沿岸带可以有自然植被或鹅卵石基质根据湖和波浪作用的类型。


巨石在沙1:本地有机细薄矩阵鹅卵石,砾石,l * 1 * ^碎片或泥浆和有机碎屑
巨石在沙1:本地有机细薄矩阵鹅卵石,砾石,l * 1 * ^碎片或泥浆和有机碎屑
图3的例子在滨海区生境异质性和斜率的影响底物组成。从Moeller再保险(1978)镜面湖的水生植物:植被结构和季节性生物量动态的研究,博士论文,212页。伊萨卡康奈尔大学纽约:。
营养动力学的滨海区
的营养来源输入碎屑(死亡的有机物)和相关的养分输入到滨海带移置(来自陆地源)或原地(水源)。移置来源包括地下水、降水、河流输入、陆生植物垃圾,从水土流失和材料。营养也可以运输到滨海带动物移动之间的陆地和沿岸带(即食品资源。哺乳动物、两栖动物,水禽,或如海狸,等等)或作为食物资源(即。小型哺乳动物,鱼类等水生生物。原地的营养来源来自死亡的水生生物(植物和动物),和分泌,排泄,排泄从活的动物和植物。
碎屑的分布影响的可用性为生物吸收溶解有机物和营养物质。碎屑沉积的深水层可能会永久失去了从滨海食物网,而碎屑沉积在沿岸带可以促进内部加载(即溶解有机物和营养。、磷和氮)为主要和次要生产。大部分的能源驱动生态系统的新陈代谢来自外来的或原地碎屑,和浅湖大百分比的滨海地区净碎屑有机质的沉积速率总是大于更深的湖泊。
保留能力的沿岸带水的保留时间,营养,和碎屑的影响大小和滨海带的配置。深湖水保留时间更长(高达数百或数千年)与浅水湖泊(通常是不到一年)和远洋区

高的景观
低景观
图4湖泊景观。水文连通性范围从孤立的湖泊所连接的大河流。Magnuson JJ, Kratz TK,本森BJ (eds)(2006)长期动态的湖泊景观:长期生态研究温带北部湖泊,51页。牛津大学,英国牛津大学出版社。
高的景观
低景观
图4湖泊景观。水文连通性范围从孤立的湖泊所连接的大河流。Magnuson JJ, Kratz TK,本森BJ (eds)(2006)长期动态的湖泊景观:长期生态研究温带北部湖泊,51页。牛津大学,英国牛津大学出版社。
已经不再保留时间与滨海区。水是保留的时间在沿岸带湖内的营养动力学的影响。时间越长对水通过滨海区,越大的营养量将由滨海带植物和动物。
虽然深湖泊更有保留时间的水,他们通常有一个小沿岸带不断失去碎屑和营养物质通过metalim-nion pro-fundal区随着碎屑。在一些分层湖泊,一半的总磷可以输给了深水层(profun-dal区)在夏季和返回的只是部分混合的湖在春天和秋天。浅水湖泊,另一方面,没有这个常数养分损失,因为他们有一个大比例的变温层体积接触到湖底。因此在浅水湖泊,营养物质循环在沿岸带更大的速度和更少的深水层发生损失。
滨海区也因此被描述为一个“代谢筛”或“陷阱”,因为它的应变能力传入的水分和养分,才向远洋深水层。在许多情况下,大多数的溶解有机物和营养,不习惯在沿岸带最终将失去在深水层沉积和埋葬。
沿岸带的主要营养成分有很多营养和矿物质(硅、钙、铁、锰、硫等)影响的化学和生物过程的类型发生在沿海地带。例如,高浓度的离子,如钙和镁增加湖泊的缓冲能力。铁和锰结合磷(通常是最营养限制初级生产)在有氧条件下生物吸收使其不可用。钙是用蜗牛壳或外骨骼维护和其他无脊椎动物,而海绵和硅藻硅骨针和测试建设要求。
滨海生物群不仅需要营养和矿物质,但反过来生物如细菌、大型植物、底栖无脊椎动物,benthivorous鱼改变可用性和沿岸带内的营养成分。自养和异养细菌可以使用和产生许多不同的营养物质和气体包括氧气、二氧化碳、铁、几个氮和硫产品,和甲烷,这取决于有氧或无氧条件。大型植物修改滨海带的化学成分通过改变氧气和二氧化碳浓度和pH值在周围沉积物和上覆水。底栖无脊椎动物和鱼类增加养分释放,如磷,通过沉积物再悬浮。
滨海的生物群区
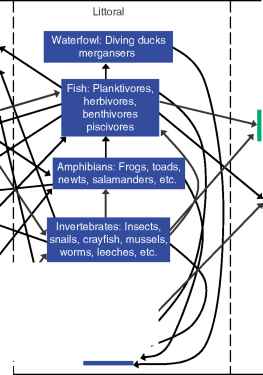
生物群的滨海区包括永久和瞬态物种(图5)。
瞬态物种——那些进出的滨海区周围的陆地生态系统和远洋区创建滨海区与周围环境之间的联系。生物群和相关的联系都是在这一节中讨论。
为便于构建食物链,生物在图5中分组被陆地或水生。然而,某些物种在每组实际上属于陆地生态系统和滨海区(即。、两栖动物和水禽)或在沿海和远洋区(即。浮游动物和鱼类)。重要的是,许多物种,包括其中一些下面讨论,使用多个食品资源和区域内的湖泊,可以有级联效应在陆地、沿海和远洋食物网。
图5沿岸带与陆地生态系统之间的联系以及远洋区。箭头表示能量流。只有与滨海带的交互显示。有陆地上的生物之间的相互作用和在深海区,并不在这个图。
陆地
水鸟:涉猎鸭子野鸭、天鹅、鹅其他鸟类:海鸥,燕鸥,鹰
哺乳动物:水獭、海狸、浣熊、老鼠等。
爬行动物:蛇、海龟
昆虫:豆娘,蜻蜓,蜉蝣,等等。
水鸟:涉猎鸭子野鸭、天鹅、鹅其他鸟类:海鸥,燕鸥,鹰
哺乳动物:水獭、海狸、浣熊、老鼠等。
昆虫:豆娘,蜻蜓,蜉蝣,等等。
远洋
鱼:食planktivores
浮游动物
远洋
鱼:食planktivores
浮游动物
一个 |
|
大型植物/固着生物1 |
|
细菌 细菌细菌生产高达120倍的滨海区比深海区。细菌的一个主要生物组件允许沿岸带作为代谢筛。细菌在沿岸带的主要功能是分解移置和原地有机材料。碎屑随着细菌的过程,不同的营养物质和气体,如微粒和溶解有机质、氮、磷、甲烷和硫等,生产和在许多情况下可用其他生物群在滨海区。与大型湖泊,浅的沿海地区将增加细菌的新陈代谢和更快的碎屑处理。 固着生物固着生物是自养和异养微生物的混合嵌入在一个矩阵的有机碎屑(指“参见”部分)。固着生物覆盖大多数水下基板,从砂岩石大型植物。固着生物代谢的重要性在整个湖规模受到沿岸带的形态测量学和基质特征。在贫营养湖泊,甚至那些有几个大型植物固着生物的生长,固着生物可以整个湖水初级生产的一个重要组成部分。例如,在一个贫营养湖,湖的沿岸带由只有15%,但固着生物占70 - 85%的湖泊初级生产。在湖泊富营养化然而,浮游植物更加丰富、阴影的浮游植物减少了固着生物和大型植物丰度。固着生物是一种常见的无脊椎动物和两栖动物的食物来源。 大型植物大型植物需要特定的底物类型茁壮成长,他们的经济增长提供了一个独特的其他生物的栖息地(指“参见”部分)。大型植物生长最好的混合砂和淤泥,并经常发现地区的地下水上涌。大型植物一旦确立在沿岸带他们通过减少修改小气候波能量和热梯度的创建,可以防止水混合。raybet雷竞技最新这些条件促进粒子沉降。小气候修改的程度取决于沉积物的特征结构raybet雷竞技最新,营养的可用性,并通过泥沙扩散的氧气。大型植物是不可或缺的养分循环滨海带的源和汇的营养。传统上,湖泊学家认为大型植物的营养来源,因为它们可能包含从缺氧沉积物营养,然后放回水柱在衰老。其他人发现,营养从沉积物或周围水体的植物主要是保留植物在植物腐烂。 除了他们的角色在养分循环,macro-phytes提供生物如细菌的重要栖息地,固着生物、浮游动物、无脊椎动物、两栖动物、鱼类和水禽。无脊椎动物和小鱼用大型植物的栖息地躲避捕食无脊椎动物(例如,蜻蜓或豆娘仙女),鱼(例如,该河)和两栖动物,繁殖的地方。对于许多无脊椎动物(如昆虫、甲壳类动物)和脊椎动物(例如,水禽,驼鹿),大型植物是一个主要的食物来源。 无脊椎动物无脊椎动物是非常多样的,包括:浮游动物、小龙虾、昆虫、蠕虫和水蛭。无脊椎动物生活在湖泊的底部被称为底栖动物,更加丰富和多样化的滨海区比其他湖泊区。因此丰富的底栖动物比浮游动物湖大小负相关。然而,绝对的多样性和丰富的底栖动物随湖的大小。无脊椎动物多样性也是正相关的生境复杂性、大型植物丰富,电导率和流连接的存在。 栖息地的可用性在沿岸带影响的无脊椎动物的类型将殖民(图2)。例如,蜉蝣(蜉蝣)和plecoptera(石蝇)通常更喜欢有更高的波浪作用的底物和粗基质,而轻易打扰chironomids是殖民的细颗粒泥沙(蚊幼虫),双壳类(蛤),和oli-gochaetes(蠕虫)。衬底、大型植物丰度和碎屑的三个主要因素控制无脊椎动物的多样性和分布,但水深,波接触,和水清晰,前三个主要因素(影响)可能也因此影响无脊椎动物丰度和分布。 鱼 像无脊椎动物多样性,鱼与湖的大小和栖息地的多样性是正相关的复杂性。不同滨海带的使用鱼栖息地也常常不同季节和年龄或鱼的大小。正如上面提到的,大型植物避难所和食肉鱼类的狩猎场。一些鱼类(例如,如spp。)也使用大型植物作为产卵基质。鹅卵石基质内其他鱼类产卵,和一些使用鹅卵石作为避难所(例如,饭桶,镖鲈,少年淡水鳕)。 鱼通常是由他们的主要食物来源分类。食鱼主要吃其他鱼类,planktivores消耗浮游生物,benthivores以底栖动物,食草动物吃植物和固着生物。一些鱼类可能会改变他们吃成熟到成年。例如,许多鱼类吃浮游生物作为一个少年和较小的鱼作为一个成年人。甚至成年鱼可能有非常广泛的饮食,因为他们之间移动沿海和远洋区(图5)。 陆地——滨海链接是不可能分离的过程发生在一个湖周围流域甚至空中分水岭。周围许多生物资源和能源之间的分水岭,滨海区。沿岸带的食物资源是一种重要的能量来源对许多陆地和semi-terrestrial生物。鱼,例如,被许多不同的陆地和两栖物种包括水禽,鹰派,鹭、白鹭,哺乳动物,爬行动物,和人类。水生无脊椎动物中发现沿岸带提供了一个重要的陆地和semi-terrestrial生物的蛋白质来源。鸭子的生殖成功密切相关的可用性chironomids新兴从底栖生物和其它昆虫幼虫形式。水禽,如鹅,以水生植物为食,可以去除50%的站在一些地区的大型植物。 食物资源沿岸陆地生态系统也很重要的物种。根据时间和鱼类,一半的食物消费可以移置昆虫和小型哺乳动物。另一种类型的资源,从土地到滨海带大伍迪碎片用海狸建造他们的小屋。使用的木质碎片海狸还提供了许多鱼类的栖息地。最后,瞬态生物如水鸟、水貂、水獭、海狸、麝鼠,蛇,和海龟,其中,营养的滨海区通过进食和排泄和排泄。 河岸栖息地是另一个资源,对使用滨海带的物种很重要。例如,蛇和龟晒太阳日志和岩石上发现沿着海岸线。水禽和一些哺乳动物使用低洼海岸栖息地筑巢,而鹰和一些潜水鸭为筑巢区使用树木周围的湖泊。鹰派也用湖周围的树栖木上寻找食物。河岸生境对两栖动物很重要(例如,蝾螈青蛙)在不同的时间在他们的生活。 营养级联,食物网交互强烈改变三个或更多的丰富营养水平——是有据可查的远洋和沿海地区的湖泊。他们还穿过littoralland界面(图6)。植物有鱼的池塘附近会有更多从传粉者访问比池塘附近的植物没有鱼。这是因为在池塘的鱼,幼虫蜻蜓是减少鱼捕食,因此大量的成年蜻蜓也降低了。成年蜻蜓有直接和间接影响昆虫传粉者。他们直接捕食传粉者和间接减少传粉者访问仅仅昙花一现。 沿岸——远洋链接沿海和远洋区也密切相关,尤其是一昼夜的水平移动的浮游动物,鱼的动作。浮游动物有时水平上升到30米区域之间每天两次。浮游动物,通常驻留在深海区将进入大型植物栖息地白天避免海洋掠食者,如Chaoborus(幻影蚊幼虫)和视觉喂养planktivores像小型鱼类。在某些湖泊,这个运动可以通过减少福利浮游动物死亡率鱼捕食,粮食供应在沿岸带(有些浮游动物可以成为滨海带的浏览器而被滤食动物在远洋区),和增强的增长。捕食planktivores往往减少了迁移到沿海区,但在某些湖泊、海滨无脊椎动物(如蜻蜓幼虫)构成实质性威胁沿岸带内的捕食。因此,浮游动物运动取决于复杂的交互发生在远洋和沿海地区,湖泊之间的不同。 鱼的动作也连接沿海和远洋区。依赖鱼鱼在沿海生产不同的类型,与planktivores benthivores,甚至食鱼依靠沿海粮食生产在一定程度上(图7)。鱼通常归类为远洋planktivores可以高达30%的能源来自滨海区,而鱼分为食鱼有时得到几乎所有的精力,至少间接(例如,从其他鱼,消耗littoral-derived食物),从滨海带(图7)。没有沿岸带,很多鱼类的生产,包括鱼,可能很少冒险进入滨海区,将大幅下降。 传粉者  鱼的年代 鱼 图6的例子一个营养级联连接滨海带的陆地生态系统。鱼减少大量的蜻蜓,导致增加传粉者,从而促进陆生植物的繁殖。实箭头表示直接的相互作用;虚线箭头表示间接交互。+积极互动,-代表了负面的互动。修改从骑士等等。(2005)在生态系统营养级联。自然437:880 - 883。 100 - 1 图7的范围依赖不同类型的鱼类对滨海带的资源。修改从Vadeboncoeur等等。(2005)的影响multi-chain杂食性营养控制的力量湖泊。生态系统8:682 - 693。 沿岸带的人为影响人类获得许多生态系统货物(例如,收获鱼类和水禽)和服务(例如,水净化,水的供应从沿海)区域的湖泊。反过来,人类有巨大的影响在沿岸带的结构和功能。首先,增加营养加载等活动日志,eutrophi-cation农业和发展原因。富营养化导致初级生产增加许多湖泊沿岸带,这可能会导致不良的海藻,以及增加不良鱼和浑浊的水。其次,人工中介传播入侵物种(如斑马贻贝,生锈的小龙虾,一轮鰕虎鱼)改变养分循环和食物网组成的滨海区,导致变化,通常是不受欢迎的人。 第三,ios版雷竞技官网入口 燃烧在工业和汽车的原因酸沉积和气候变raybet雷竞技最新化。在许多情况下,湖泊的酸化导致大型植物丰度和多样性降低,无脊椎动物,鱼,同时增加绿色丝状藻类生产,所有的层叠效果,通过食物网。酸化还会导致金属的释放有毒鱼类,如铝和汞。这些金属的生物蓄积在鱼类,然后使鱼对人类有害。raybet雷竞技最新气候变化预计将导致变暖湖水域,在世界的许多地方,将减少径流,增加水的居留时间,降低水位,增加蒸发。因此许多离子浓度增加,导致养分变化和碎屑的可用性以及中小学在沿岸带生产。气候变暖也可能导致向极范围扩大许多沿海的物种,进一步改变食物网动力学。 最后,在水位波动往往增加了灌溉和大坝。在高水位、洪水和河岸栖息地发生侵蚀。这就增加了溶解有机质的输入和浊度的滨海区。另一方面,水减少正面和负面影响滨海区,根据湖盆形态测量学。陡峭的滨海区域不像浅倾斜的影响的。在低水位大型植物减少,桑迪/细粒栖息地百分比的增加,底栖无脊椎动物多样性和大量减少和鱼类保护区和产卵栖息地可以减少。 所有这些沿岸带的反应人为影响受到沿岸带的结构和功能以及沿岸带和陆地生态系统之间的交互和远洋深水区。湖响应的程度或湖的时间能抵抗被上述humanmediated压力的影响取决于滨海带的大小,在景观湖的位置,不同栖息地的丰度和分布在滨海区,和不同的生物群,区内。因此,整个湖泊沿岸带是重要的功能以及对人类的反应整个湖。 参见:底栖无脊椎动物、湖泊和水库;气候变化对湖泊的影响;raybet雷竞技最新富营养化的湖泊和水库;浅湖泊和池塘。 进一步的阅读伯克斯r . L,洛奇d M, Jeppesen E,和Lauridsen t L(2002)一昼夜的浮游动物的水平迁移:存在于这个沿海的成本与效益。淡水生物学47:343 - 365。 Gasith和Gafny年代(1990)sturucture水位波动的影响和滨海带的功能。:Tilzer M C M和Serruya (eds)。大湖:生态结构和功能,156 - 171页。纽约:Srpinger。 Jeppesen E,桑德迦米、桑德迦米,Christoffersen K (eds)(1998)在湖泊SubmergedMacrophytes的构建作用。纽约:施普林格。 Kalff J(2002)湖沼学:内陆水生态系统。上台北:新世纪。 骑士t M,真品M . W,追逐j . M, et al。(2005)在生态系统营养级联。自然437:880 - 883。 把g . E (ed)(1985)水生生态生态系统方法。纽约:施普林格。 洛奇D M, Barko j . W,流浪者D, et al。(1988)空间异质性和栖息地在湖社区的交互。:木匠s R (ed)湖社区的复杂的相互作用。纽约:施普林格。 Magnuson J . J Kratz t·K,本森b J (eds)(2006)长期动态的湖泊景观:长期生态研究温带北部湖泊。英国牛津:牛津大学出版社。 雷诺兹奥沙利文p E和c . S (eds)(2004)湖泊手册。英国牛津大学:布莱克韦尔科学。 Pieczynska E(1993)碎屑和营养动力学的海岸区湖泊:复习一下。Hydrobiologia 251: 49-58。 雅伯M(1998)浅水湖泊的生态。纽约:查普曼和大厅。 Vadeboncoeur Y,麦肯k . S VanderZanden m J,拉斯穆森J . B(2005)的影响multi-chain杂食性在湖泊营养控制的力量。生态系统8:682 - 693。 韦瑟m和詹姆斯·m·R (2001) macro-invertebrates分布与物理和生物变量九新西兰湖泊沿岸带的。Hydrobiologia 462: 115 - 129。 吉姆r G(1989)地面水达到接口:代谢和limno-logical监管机构。国际理论与应用协会湖沼学程序24:6日到24日。 吉姆r G(1989)湖沼学:湖和河流生态系统。纽约:学术出版社。 |
|
继续阅读:底栖无脊椎动物的湖泊和水库
这篇文章有用吗?
推荐项目



读者的问题
-
Senait5天前
- 回复
-
厄休拉5天前
- 回复
-
Kerstin5天前
- 回复
-
乔5天前
- 回复
-
迈克尔5天前
- 回复
-
坎贝尔·斯图尔特5天前
- 回复
-
利兰5天前
- 回复
-
梅丽莎5天前
- 回复
-
亨利5天前
- 回复
-
asmarina5天前
- 回复
-
艾丽西亚乔丹5天前
- 回复
-
马丁•齐默5天前
- 回复
-
senay5天前
- 回复
-
套1个月前
- 回复
-
riia1个月前
- 回复
-
省略1个月前
- 回复
-
adaldrida扮演1个月前
- 回复
-
克里斯汀1个月前
- 回复
-
穆雷1个月前
- 回复
-
修纳人赖利1个月前
- 回复
-
ZULA1个月前
- 回复
-
马太福音1个月前
- 回复
-
丽娜2个月前
- 回复
-
米蕾tesfalem2个月前
- 回复
-
桑德拉2个月前
- 回复
-
格雷戈里奥3个月前
- 回复
-
克劳迪亚3个月前
- 回复
-
汤姆3个月前
- 回复
-
格罗弗3个月前
- 回复
-
elisa3个月前
- 回复
-
查尔斯3个月前
- 回复
-
莫斯科3个月前
- 回复
-
菲利普3个月前
- 回复
-
卢卡3个月前
- 回复
-
yonatan neftalem3个月前
- 回复
-
苏珊3个月前
- 回复
-
佩特拉3个月前
- 回复
-
patryk3个月前
- 回复
-
弗里达·贝鲁奇3个月前
- 回复
-
bonacata4个月前
- 回复
-
kieron4个月前
- 回复
-
贝丝4个月前
- 回复
-
汤姆4个月前
- 回复
-
安东尼·安德森4个月前
- 回复
-
斯文4个月前
- 回复
-
segan4个月前
- 回复
-
厄休拉维斯4个月前
- 回复
-
科妮莉亚4个月前
- 回复
-
梅丽莎4个月前
- 回复
-
埃罗4个月前
- 回复
-
弗罗多sandyman4个月前
- 回复
-
艾巴勒莫4个月前
- 回复
-
玛蒂尔达4个月前
- 回复
-
米歇尔·黑4个月前
- 回复
-
维托4个月前
- 回复
-
阿塞尼奥。4个月前
- 回复
-
paola4个月前
- 回复
-
亚历山大4个月前
- 回复
-
利比4个月前
- 回复
-
kerttu4个月前
- 回复
-
保罗4个月前
- 回复
-
迈克尔4个月前
- 回复
-
taimi4个月前
- 回复
-
弗兰基4个月前
- 回复
-
金发女孩4个月前
- 回复
-
托尔斯滕4个月前
- 回复
-
kaija4个月前
- 回复
-
桑德拉4个月前
- 回复
-
克里斯蒂娜5个月前
- 回复
-
西蒙5个月前
- 回复
-
MELAMPUS5个月前
- 回复
-
玛丽娜5个月前
- 回复
-
MIIKKA5个月前
- 回复
-
丹妮拉嘉宝5个月前
- 回复
-
尼可莱塔5个月前
- 回复
-
威廉5个月前
- 回复
-
kacper5个月前
- 回复
-
futsum5个月前
- 回复
-
赛义德·迪克森5个月前
- 回复
-
tiblets阿斯马拉5个月前
- 回复
-
罗宾5个月前
- 回复
-
爱里5个月前
- 回复
-
维多利亚5个月前
- 回复
-
鲁道夫6个月前
- 回复
-
jonna做6个月前
- 回复
-
法尔科6个月前
- 回复
-
布莱恩6个月前
- 回复
-
amalda6个月前
- 回复
-
1月6个月前
- 回复
-
恩颐投资6个月前
- 回复
-
BRHANE6个月前
- 回复
-
詹姆斯6个月前
- 回复
-
Ottavio6个月前
- 回复
-
金发女孩6个月前
- 回复
-
欧文7个月前
- 回复
-
玛丽7个月前
- 回复
-
法尔科Took-Took7个月前
- 回复
-
terzo花茎甘蓝7个月前
- 回复
-
pamphila7个月前
- 回复
-
柯拉7个月前
- 回复
-
VANNA吗哪7个月前
- 回复
-
黑兹尔7个月前
- 回复
-
埃里克7个月前
- 回复
-
阿齐兹7个月前
- 回复
-
苏里士满7个月前
- 回复
-
abrha7个月前
- 回复
-
马库斯Jaeger7个月前
- 回复
-
尤金尼亚7个月前
- 回复
-
HABTE7个月前
- 回复
-
詹姆斯7个月前
- 回复
-
玛丽亚7个月前
- 回复
-
艾伦negassi8个月前
- 回复
-
安妮8个月前
- 回复
-
Tytti8个月前
- 回复
-
temesgen rezene8个月前
- 回复
-
赛斯布朗8个月前
- 回复
-
詹姆斯Chilson8个月前
- 回复
-
Madoc8个月前
- 回复
-
艾达8个月前
- 回复
-
塞拉姆8个月前
- 回复
-
贝基8个月前
- 回复
-
Berylla8个月前
- 回复
-
曼迪Ehrlichmann8个月前
- 回复
-
mirella8个月前
- 回复
-
亚莉克莎8个月前
- 回复
-
斯蒂芬8个月前
- 回复
-
celendine8个月前
- 回复
-
侍从8个月前
- 回复
-
温迪8个月前
- 回复
-
比利8个月前
- 回复
-
蒂利8个月前
- 回复
-
雨果8个月前
- 回复
-
patrycja格兰特8个月前
- 回复
-
塞布丽娜施瓦兹8个月前
- 回复
-
许多tuukkanen8个月前
- 回复
-
凯瑟琳的河流11个月前
- 回复
-
银白杨1年前
- 回复
-
马里诺1年前
- 回复
-
凡妮莎1年前
- 回复
-
斯蒂格Koskinen1年前
- 回复
-
克劳斯·舒尔茨1年前
- 回复
-
莫丽罗素1年前
- 回复
-
Tekle1年前
- 回复
-
哈姆萨1年前
- 回复
-
PETROS AMANUEL1年前
- 回复
-
罗纳德·阿什利1年前
- 回复
-
olavi1年前
- 回复
-
萝珊1年前
- 回复
-
antje1年前
- 回复
-
阿卜杜拉·克里斯蒂1年前
- 回复
-
quarantino1年前
- 回复
-
vilho1年前
- 回复
-
苏珊·埃尔南德斯1年前
- 回复
-
uta1年前
- 回复
-
Minto1年前
- 回复
-
Demet1年前
- 回复
-
kisanet1年前
- 回复
-
马太福音1年前
- 回复
-
celendine1年前
- 回复
-
ADONAY1年前
- 回复
-
CADEN1年前
- 回复