抗微生物群落的韧性和冗余
史蒂芬·d·埃里森* +和詹妮弗·b·h·她*
尽管人们普遍认为植物群落组成是预测利率的关键生态系统过程面对的全球变化,微生物群落组成往往是忽略了在生态系统建模。为了解决这个问题,我们回顾最近的实验和评估是否耐药微生物群落组成,有弹性,或者功能冗余以应对四个不同的干扰。我们发现大多数微生物组的成分敏感,不能马上适应干扰,不管分类组的宽度或干扰的类型。其他研究表明,组合的变化往往与生态系统的变化过程。因此,微生物群落的变化由于扰动可能直接影响生态系统过程。基于这些关系,我们提出一个简单的框架将微生物群落组成生态系统过程模型。我们得出结论,这种努力将会从中受益更多的微生物之间的链接发展史上的一次经验数据,生理特征,和扰动的响应。这些关系将决定容易微生物群落组成可以用来预测生态系统过程对全球变化的响应。
部门“生态学和进化生物学和地球系统科学,加州大学欧文分校CA 92697。
150 /史蒂文·d·埃里森和詹妮弗·b·h·她
最近的植物和动物物种的灭绝率促使生态学家认为生物多样性丧失的后果。超出了伦理和审美原因保护,生物多样性经济价值的生态系统货物和服务供应人类社会赖以(埃尔利希和欧利希,1992;每天,1997)。尽管大多数生物多样性和保护的研究集中在大型生物的价值和重要性,非常大量的微生物,给予他们一个主要角色在提供生态系统服务,如水净化和土壤肥力。细菌和古菌仅包含大部分的总氮(N)和磷(P)和一半的碳(C)存储在生物体(惠特曼et al ., 1998),和微生物的代谢机械驱动各种生态系统的过程。事实上,微生物进行分解和催化的大部分重要的转换C, N,硫和P周期。
尽管他们对生态系统的功能的重要性,微生物很少明确地考虑各个生态系统或全球流程模型。除了方法论的障碍,这一差距的主要原因是他们的压倒性的多样性。估计土壤微生物多样性的范围从几千到一百万几克土壤中微生物“物种”(Torsvik和0 vreas, 2002;甘斯et al ., 2005),以及这种多样性与生态系统过程通常是未知的(Torsvik等,2002;克劳福德等,2005;阿扎姆和极糟,2007)。此外,它是不可行,评估和跟踪每个微生物分类单元在一个生态系统,更不用说甚至包括一小部分这些类群的生态系统模型。
因为这些障碍,常常“黑匣子”微生物生态系统模型。换句话说,微生物被埋在方程结构动力学常数和响应函数和“简化得面目全非”(丢弃,2001)。结果,丰度、多样性和微生物的相互作用通常被认为是不重要的生态过程,尤其是在陆地生态系统模型。(大量的海洋生态系统模型包括各种浮游植物组织(例如,摩尔et al ., 2002;Salihoglu和霍夫曼,2007)]。
与微生物相比,人们普遍认为植物生物多样性(包括丰富性和组成)影响陆地生态系统进程(Tilman et al ., 1997;赫克托耳et al ., 1999;Spehn et al ., 2005)和影响生态系统对干扰的反应,如二氧化碳和N(如。帝国et al。(2001)]。植物群落组成通常是纳入大规模模型通过使用官能团,基于植物性状(Tilman et al ., 1997;帝国et al ., 2007)。全球变化生态系统模型(Haxeltine普伦蒂斯,1996;默克罗夫特et al ., 2001),陆地生物圈(Foley et al ., 1996),或全球气候(考克斯et al ., 2000;raybet雷竞技最新希金斯和施耐德,2005)通常包含5 - 10植物功能群来提高模型的预测。
尽管如此,研究人员呼吁更好的理解植物的功能特征来预测生态系统进程在不断变化的环境条件下(Diaz et al ., 2004;阿恩特,2006;麦吉尔et al ., 2006)。
鉴于微生物在生态系统过程的核心作用,我们认为微生物成分可能至少同样重要植物成分为构建预测生态系统模型。在这里,我们讨论两个主要障碍包括微生物在生态系统模型。第一个是一般认为微生物成分并不重要生态系统过程。第二,微生物成分可能过于多元化的模型。解决第一个障碍,我们概述三个条件,需要对微生物组成重要生态系统过程的变化。然后我们回顾最近的研究来评估特定的微生物群体是否可能或多或少受到特定的干扰。我们解决第二个障碍提出了微生物过程的一个简单的模型,包含社区组成的信息。模型是用来说明实证数据可以用来预测微生物过程扰动下,即使是相对多元化的社区。因为它不是可行的添加一个参数为每个微生物种群在一个社区,我们认为当粗微生物组成、信息的相对丰度等几个关键演化支在一个官能团,可以帮助减少不确定性的预测生态系统过程。
黑盒内的一个框架
丢弃(2001)指出,生态系统黑盒模型使两个隐式的假设:微生物过程可以表示在一系列环境条件与一个数学函数,而微生物过程永远不会受到任何微生物的丰富。这些假设是隐式的,因为没有明确主要生态系统模型包括参数代表微生物群落组成的组件。在最好的情况下,一些模型包括总微生物生物量作为参数(如。,哈特和Kinzig(1993)),但许多广泛使用的模型如世纪(帕顿等,1987),梅尔(多个元素限制)(Rastetter et al ., 1997),和TEM(陆地生态系统模型)(McGuire et al ., 1993)包含参数相关的微生物过程但不是微生物群落本身。海洋biogeochem-istry模型才刚刚开始将显式参数捕捉浮游生物群落组成(摩尔et al ., 2002;Le Quere et al ., 2005;Salihoglu和霍夫曼,2007)。
这些模型隐式假设社区组成的变化不会影响生态系统过程,因为没有机制
152 /史蒂文·d·埃里森和珍妮弗•b . h .她为这些变化影响模型预测。这种假设可能有效如果耐药微生物组成,弹性和/或功能冗余(这些条款和其他人在表8.1中定义)。抗微生物组合是如果是相似的在各种环境条件句话说,很难从一个扰乱初始状态。它是有弹性的,如果当成分改变,恢复迅速,无论是增长或由生理或遗传适应。最后,在应对新环境条件下的微生物成分可能会改变但包含功能冗余等类群,生态系统过程利率不会改变。如果这些条件不是一般适用于微生物群落,然后许多当前生态系统模型以预测利率可能会失败microbe-driven过程扰动下。
抗微生物的想法、弹性和生态功能冗余是遍布。一些微生物群体表现出高度的代谢灵活性和生理公差改变环境条件(如。梅尔et al。(2004)),这可能导致微生物群落是难以改变的。这些特征和其他与microbes-such丰度高,广泛传播,以及潜在的快速增长利率也导致微生物群落的建议会改变(Fenchel和弹性
表8.1总结本章定义中使用
术语
定义
官能团的功能冗余
功能相似
微生物组成
微生物的分类单元的弹性
电阻
生理特征
生理响应曲线
所有生物体直接导致特定功能过程的速率在一个生态系统微生物分类单元的能力进行一个过程以同样的速度作为另一个相同的环境条件下的能力两个微生物群落进行功能过程以相似的速度,无论差异成分丰富,类群的相对多度,系统结构组合的一组系统相关微生物的微生物组成后回到原来的成分干扰微生物的组成不变的脸干扰微生物的生理特点决定了贡献分类单元功能流程
函数描述之间的关系过程微生物分类单元和扰动强度的贡献
芬利,2004;梅耶,1994)。此外,通过水平基因转移可能允许快速进化适应敏感微生物的适应新的环境条件和迅速恢复原来的社区组成。微生物的极高的丰度和多样性作为参数用于功能冗余,因为很难想象,生物地球化学循环受到微生物丰度(梅耶,1994)或遗传多样性(芬利et al ., 1997)。
图8.1说明了潜在影响的干扰微生物组成和/或生态系统的过程。考虑干扰应用到一个生态系统,其中的微生物群落。抗微生物组合可能会干扰,不会改变。或者,如果社区是敏感和改变,这可能是弹性和迅速恢复到最初组成。最后,一个社区的
-
.")
- 图8.1一个示意性的干扰可以改变微生物的组成,从而影响生态系统过程与当扰动不会有这种效果(耐药微生物群落时,弹性或功能冗余)。
.")
154 /史蒂文·d·埃里森和珍妮弗•b . h .她的作文很敏感,没有弹性的利率可能生产过程类似于原始的社区社区成员在功能上是冗余的。
只有社区组成受干扰,没有弹性,功能不同的原社区做社区组成物质的变化预测生态系统过程。此外,社区是耐药的程度,有弹性,功能类似的会影响社区组成的程度关系到一个特定的过程。例如,即使微生物组成高度敏感和没有弹性的干扰,如果所有的分类单元执行一个过程非常相似率(也就是说,他们几乎是功能冗余),那么生态系统过程的预测利率将不会改善,包括微生物组成的信息。
这个概念框架并不排除这种可能性几乎没有微生物组成的变化,但大生态系统的变化过程。这个场景中建议的变化是环境变化的直接影响,成分变化的结果在其他生物如植物或微生物的生理反应,不是伴随着成分变化。后者的一个例子是一些转换等分解速度发生在更高的温度。这是见过立即组成的变化可能是负责(菲勒et al ., 2005)。
抗微生物组成
评估是否经常抵抗干扰微生物组成,我们回顾了研究实验性地暴露了微生物群落的各种干扰。我们搜索网络的科学论文包括“活*”和“社区组成”头衔,抽象,或主题的单词。此外,我们专门在《全球变化生物学》杂志上搜索全球变化实验,评估micro-bial组成。我们没有限制论文通过研究系统,但大部分研究返回这些搜索参数集中在陆地土壤。我们承认,有许多的海洋研究,我们没有在我们的搜索。干扰我们的研究仅限于二氧化碳浓缩,温度,和矿物营养施肥,浓缩与C基质(包括复杂organ-ics如肥料和杀虫剂等潜在的毒素)。我们这四个障碍的原因是典型的那些经验人为全球变化生态系统。最后,我们不打算详尽的文献检索,但代表的这些类型的研究。
我们假设研究重点和方法的差异会影响检测组分反应的可能性。因此,对于每一个研究我们记录的广度taxo-nomic集团目标(例如,细菌和真菌;methanotrophic细菌),栖息地,方法用于检查作文,和干扰,以及目标群体的组成是否改变了扰动响应。我们还记录了时间成分的干扰首次应用后评估,有关讨论下面的韧性。
论文从这个搜索包括研究目标成分大致如磷脂脂肪酸分析、目标细菌和真菌。相比之下,其他的研究检查窄功能公会内的细菌PCR-amplifying功能基因,后跟一个指纹识别技术(如变性梯度凝胶电泳或终端限制片段长度多态性)。因此,研究类群的广度差异很大,相关方法用于评价社区组成。方法本身也可能会影响是否检测成分的变化。例如,序列扩增克隆菌进行库将揭示finer-scale基因变化比指纹识别方法,目标相同的基因。最后,因为我们感兴趣的成分变化,我们的搜索排除许多研究只测量总micro-bial生物质(如。,那些了瓦尔德(1992)]。
表8.2总结了四种干扰的结果。大多数这些研究证明成分对干扰很敏感。超过80%的矿物受精(N / P / K),温度,和C修正案研究发现显著的扰动对微生物组成的影响。相比之下,高二氧化碳的影响很少被发现,只有60%的研究。
的平均长度 |
|||||
不。的 |
的百分比 |
研究(年 |
±SD) |
||
干扰 |
研究 |
研究敏感 |
耐药 |
敏感的 |
P值 |
二氧化碳的增加 |
20. |
60 |
3.3±1.9 |
3.6±1.9 |
0.78 |
N / P / K受精 |
38 |
84年 |
4.8±4.5 |
8.2±8.7 |
0.17 |
温度 |
11 |
82年 |
1.1±1.3 |
3.0±5.0 |
0.35 |
C修正案 |
41 |
83年 |
0.15±0.09 |
4.9±12.6 |
0.03 |
注意:P值报告是一个t测试的结果(假设不平等的方差)之间的研究时间研究抗性与敏感。
注意:P值报告是一个t测试的结果(假设不平等的方差)之间的研究时间研究抗性与敏感。
156 /史蒂文·d·埃里森和詹妮弗·b·h·她
这些研究并没有表明,广泛的分类群的或多或少对干扰敏感比狭窄的分类群。该模式表明,分类广度与成分变化是否检测到。或许更令人惊讶的是,没有模式表明方法影响成分变化是否检测到。此外,我们无法辨别是否特定的分类或特定官能团或多或少敏感扰动类型。总的来说,低数量的研究观察耐药微生物成分阻碍了我们这些研究中识别任何模式的能力。不过,我们可以得出这样的结论:微生物成分通常是对干扰敏感。
弹性的微生物组成
即使微生物组成受干扰、社区可能仍然是有弹性和迅速恢复predisturbance组成。许多微生物的特性,特别是细菌和古生菌,表明弹性可能是常见的。首先,许多微生物快速增长;因此,如果他们的富足是抑制扰动,有可能很快恢复。第二,许多微生物有一个高度的生理的灵活性。这是著名的紫色nonsulfur细菌,可光下缺氧条件下在有氧条件下,异养生物。因此,即使一些类群的相对多度降低最初,这些类群可能生理适应新的非生物条件随时间和返回到原来的丰度。raybet雷竞技最新最后,如果生理上的适应是不可能的,那么快速进化(通过突变或水平基因交换)可以让微生物类群适应新的环境条件,从干扰中恢复过来。所有这些参数假设富足是减少干扰,但一些微生物类群可能受益于新的条件和丰富的增加。因此,为了一些类群丰富的恢复,那些积极回应干扰也需要减少丰富社区重返原来的成分。
很少有研究明确地关注微生物组合扰动后的时间进程;相反,大部分只关注成分的敏感性。因此,我们记录的时间应用程序之间的干扰,当微生物组合评估的研究在我们的样例。如果组合是高度弹性,那么应该不太可能从扰动检测成分变化随着时间的增加。
我们比较这些研究发现的时间从最初的干扰成分敏感与耐药。通常,蒂姆的荷兰国际集团(ing)组成评估差异很大,从几小时到几十年。对于C修正案,微生物群落组成变化的研究明显长于研究没有发现变化(表8.2)。这个结果意味着有一个滞后的反应微生物群落C添加,不支持这些社区是有弹性的。矿物受精,二氧化碳升高和温度,所有研究都同样可能会发现社区组成的变化,不管自干扰。平均而言,回顾了研究检查成分干扰应用程序的几年后。因此,作为一个保守的边界、微生物成分通常是在几年内没有弹性。
当然,干扰的强度和频率应用会影响微生物组成的弹性。大部分的研究中我们回顾了继续应用扰动研究(如发生对大多数全球变化干扰),而不是一次性应用在实验的开始。例如,Enwall et al。(2007)受精和土地不肥沃的土壤图相比,自1956年以来一直保持。一般细菌的构成和ammonia-oxidizing细菌仍然不同图表类型。相比之下,鲜明的et al .(2007)应用有机和无机形式的N土壤样本和放线菌的成分相比,alpha-Proteobacteria和假单胞菌。10天后,成分之间的不同土壤的治疗方法,但91天后组成不同只在假单胞菌。相反,一些研究没有发现扰动对成分的影响可能会发现如果产生影响进行了再研究。
功能冗余的微生物组成
我们的文献调查清楚地表明,微生物群落对干扰敏感,常常不迅速恢复到原来的状态。这些反应求的问题组成的变化是否会影响生态系统流程,扰乱社会功能类似于原始社会吗?有两个原因微生物组合可能不会影响生态系统过程的变化。首先,新社区可能包含功能冗余的类群,类群在老社区。第二,新社区的类群可能相同功能不同,但是导致过程速率结合时在社区一级。
我们知道在微生物群落功能冗余?一些研究比较不同微生物功能群内的冗余程度(例如,但看到沃尔et al。(2004)和Setala和麦克莱恩(2004)]。功能冗余国,是困难的
158 /史蒂文·d·埃里森和詹妮弗·b·h·她丽斯,因为它需要执行特定的微生物种群详细知识的过程。此外,生物功能冗余的一组条件下不得在不同条件下。一般来说,我们知道一些关于功能的分布特征在微生物类群尽管多年来识别的需要(丢弃,2001)。
尽管如此,越来越多的文献开始地址是否微生物组成的更基本的问题关系到生态系统过程的任何程度和功能组的成员是否完全冗余。为了解决这个问题,需要操纵微生物成分而非生物环境的控制。这是因为任何过程的差异率可以直接归因于成分差异而不是简单的生理反应的原始社会在新的环境条件下。在植物群落组成和多样性可以直接操纵在地里播种、除草的阴谋。虽然这不是一项容易的任务,它比操纵micro-bial更可行的社区组成。
许多方法可以用来解决成分与环境对生态系统过程的影响。在一个方法中,过程率测量之前和之后的一个扰动操作但在微生物组成变化。这测量提供了一些想法的直接影响干扰过程利率独立的社区组成。丢弃和Gulledge(1998)审查研究主题从不同的栖息地平行短期微生物群落,社区环境的治疗方法和比较的过程。例如,Gulledge et al。(1997)发现,利率的甲烷氧化菌消费社区应对受精不同铵取决于社区与纸皮桦或白云杉有关针叶林。这一结果表明,菌成分的差异负责不同的生理反应。
尽管他们可能不是模仿自然的社区,生物反应器是有用的模型系统连接微生物功能群和过程。社会成分可以直接操纵测试功能的贡献不同的微生物类群。此外,社区可以摄动和监控功能和组成。例如,费尔南德斯et al。(2000)建造两个不同产甲烷反应器社区功能类似之前用葡萄糖脉冲干扰。脉冲后,社区代谢葡萄糖使用不同的路径和恢复前脉冲函数在不同的利率。作者能够属性这些差异具体组织的细菌显示不同的代谢功能。
移植实验也可以用来单独的环境和成分对过程的影响率(里德和她,2007)。如果不同的微生物群落产生不同的流程利率在一个共同的环境中,那么可以推断,负责功能的成分差异的差异。巴尔斯和费尔斯通(2005)提供了一个很好的例子的移植方法也可以用来使微生物类群之间的联系和处理利率受到干扰。他们移植土壤微生物群落在气候梯度和证明社区组成独立的气候过程影响利率。raybet雷竞技最新此外,他们使用磷脂脂肪酸数据关联过程与特定的微生物群落的成员和得出结论,硝化潜力和一氧化二氮通量可能是由革兰氏阴性细菌。
虽然不是经常,微生物组成的直接操作可以提供有用的微生物群体的功能状态的信息,尤其是当加上过程速度测量。例如,可以针对特定的分类单元从一个社区通过化学或物理手段和过程相比在社区和没有类群(桑托斯和美国华福,1981;格里菲思et al ., 2000;奥斯汀et al ., 2006)。Wertz et al。(2007)控制土壤微生物组成的连续稀释和reinnoculation无菌的缩影;他们没有发现微观结构对功能的影响。另外,社区可以人为地构造包含特定类群和建立成分和过程之间的联系(Naeem et al ., 2000)。例如,贝尔et al。(2005)表明,细菌的多样性和组成影响呼吸率水生缩影。
上面的部分文献综述表明,微生物成分往往是被干扰,不改变在一段时间恢复。此外,这些变化往往影响生态系统过程的速率,表明至少某些微生物类群在功能上是不同的。根据这些观察结果,我们提出一个广泛的框架下一节将微生物组成信息集成到生态系统过程的预测模型。
将微生物纳入模型:生理特征和过程响应曲线
随着越来越多的数据被收集在微生物组成和生态系统功能之间的关系,明确地将微生物纳入流程模型将变得越来越容易处理。事实上,类似的努力已经成功的与植物功能群和生态系统模型。然而,有一些差距微生物ecolo——之间的桥梁
160 /史蒂文·d·埃里森和珍妮弗·b·h·她依据和生态系统建模。建模不确定如何将极其多样化微生物群落聚合成一个可控的功能单元的数量。相反,微生物生态学家经常有一个贫穷的理解类型的研究,将有用的建模者在执行这聚合的步骤。在本节中,我们概述简单的微生物过程模型框架的一些结果,考虑实验可能会进一步通知预测模型。
考虑一些类群中官能团,有利于生态系统的过程。的官能团有n个类群丰度ai(单位生物量)和biomass-specific生理利率r。社区过程速率r的和产品的丰度和利率:
R = airi + - + anrn。
如果分类单元1和分类单元2有相同的生理利率ri,然后他们在功能上是冗余的,和他们的丰度可以聚合在一起:
a1r1 + a2r2 = (a1 + a2) r1。
如果社区1和2有不同的成分相同,但过程率(R1, R2),那么我们就可以将这些社区定义为功能相似,尽管他们可能包含不功能冗余的类群。
预测微生物过程将如何应对干扰,我们还需要知道每一个分类单元的生理反应干扰。假设生理响应曲线r(我)是一个线性函数的扰动强度:
m是斜率和r0生理率在安静的条件下(图8.2)。对于整个微生物群落,过程速率的函数扰动强度我会
因为它不是可行的模型单独所有类群及其响应,挑战是确定功能组的属性,这将有助于预测生态系统的响应过程扰动。未来实验研究是一个值得的目标识别的冗余水平生理特征和干扰微生物功能群内的反应。然后我们可以估计的参数m

图8.2微生物分类单元的生理响应曲线。曲线说明了分类单元的速度有助于生态系统过程扰动强度的函数。为简单起见,这个函数被认为是线性的,尽管其他形式可能为微生物类群。直线的斜率m表示生理率变化的速度和我,和r0生理率没有干扰。
扰动强度(I)
图8.2微生物分类单元的生理响应曲线。曲线说明了分类单元的速度有助于生态系统过程扰动强度的函数。为简单起见,这个函数被认为是线性的,尽管其他形式可能为微生物类群。直线的斜率m表示生理率变化的速度和我,和r0生理率没有干扰。
和r0无需测量每一个分类单元。例如,如果之间有通信发展史和生理特征(r0),或发展史和功能之间的反应(m),然后我们可以使用系统发育信息来估计参数。鉴于微生物组成与系统发育标记通常是评估,这些信息可以用来预测功能组的成员如何影响生态系统过程。
作为这种方法的例子,考虑图8.3中的官能团。所有类群的进化枝顶部执行功能过程在一个较高的率(用大圆圈),和那些在底部进化枝执行过程在一个更低的利率(用小圆圈)。如果总丰度这两个演化支保持不变,然后组成的变化在两个演化支在功能上并不重要,因为类群演化支内执行函数以同样的速度。在模型中更
162 /史蒂文·d·埃里森和詹妮弗·b·h·她
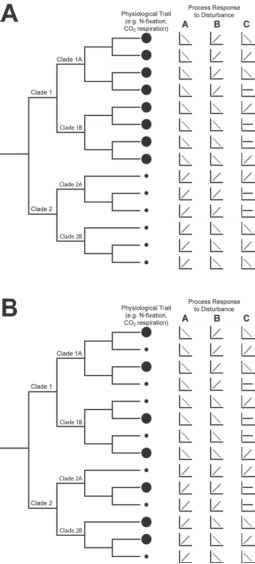
图8.3比较程式化的说明生理特征和过程的系统发育关系反应分类单元之间的干扰。不同的生理特征是由不同大小的圆圈(见文本进一步解释)。,生理特征值与系统发育相似,而在B,生理特征值是随机分布的类群之一。三个不同的干扰(A、B和C)生产过程反应的类群,也是系统相关(A和B)干扰或随机分布在类群(扰动C)。
图8.3比较程式化的说明生理特征和过程的系统发育关系反应分类单元之间的干扰。不同的生理特征是由不同大小的圆圈(见文本进一步解释)。,生理特征值与系统发育相似,而在B,生理特征值是随机分布的类群之一。三个不同的干扰(A、B和C)生产过程反应的类群,也是系统相关(A和B)干扰或随机分布在类群(扰动C)。
一般来说,这些演化支定义水平的聚合等一个安静的社区贡献过程R
R = ac1r0c1 + ac2r0c2 ac1和ac2带来总结的所有类群的演化支1和2 r0c1和r0c2生理类群的演化支1和2,分别。
与图8.3的生理特征,干扰的反应也可能与发展史。例如,内类群进化枝1所有回应同样的扰动,一样内类群进化枝2。因此,必须考虑这两种演化支的丰度来预测整个过程速率R在干扰:
R (I) = acl(制程x I + r0d) + ac2 (mc2 x + r0c2),在mc1 mc2代表的生理响应曲线的斜坡演化支1和2,分别。我们注意到之前必须确定扰动下的丰度的计算新流程速度,除非他们是已知的与扰动是稳定的。
建模的目的,这将是有用的了解生理率之间的关系,扰动响应和类群之间的系统发育地位。然后,利率的预测过程可以估计基于相对广泛的系统组织的丰度。然而,目前我们缺乏必要的经验证据来确定哪些微生物过程显示phy-logeny之间强大的通信和生理学。虽然这个话题在微生物生态学文献中已获得了高度的关注(Andren Balandreau, 1999;巴尔斯et al ., 2002;Nannipieri et al ., 2003),很少有研究测试了令人信服的联系。
干扰B在图8.3中说明情况下,生理反应曲线系统分组的subclades(表示1 a、1 B、2和2 B)。一般模型,我们将计算过程速率
R (I) = adA (mdA x 1 + r0c1) + ac1B (mc1B x 1 + r0d) + ac2A人员x 1 + r0c2) + ac2B (mc2B x 1 + r0c2)。
这个模型比之前模型复杂得多,因为干扰subclade一级反应的异质性。在一个复杂的微生物群落可能出现这种情况但仍是驯良的建模如果异质性相对较低,研究的很透彻了。例如,Horz et al。(2005)发现,两种不同的演化支
164 /史蒂文·d·埃里森和詹妮弗·b·h·她氧化菌对模拟的全球变化反应不同。最坏的情况是,分类单元的响应扰动是随机分布在种系发生的扰动C在图8.3所示。在这种情况下,整个过程的计算速度要求每个分类单元的丰度。类似的挑战会出现功能性特征本身并不是过去的决定(图8.3 b)。
克服的挑战的一个有前途的工具集这些“最坏”的情景是宏基因组或metatranscriptomic方法。例如,环境基因标签(废气温度)有可能被用作代表生理利率或扰动响应在整个社区,即使这些特征无关phy-logeny (Tringe et al ., 2005)。用这种技术,基因的丰度特定于特定环境流程(如磷酸代谢)可以从社区宏基因组数据和建模过程中使用的利率。这种方法的另一个优点是,可以检查多个废气温度和流程在一个示例中,而不是构建单独为每个不同的功能基因克隆库。
在我们的模型中,当改变组成生态系统过程的重要吗?改变一个分类单元的数量会影响处理速度,除非其他类群的丰度也发生了变化进行补偿。在安静的社区里,变化到另一个分类单元(具有类似生物质能)会影响社区过程速度如果两组有不同的r0(生理)值。干扰,如果类群组成不同的m(扰动响应)值的变化丰富(即使他们有相同的r0值)。虽然我们的简单的模型只考虑总丰度(即。,生物量中)不同的类群,我们注意体型分布在分类群的变化也可能影响生态系统过程利率所显示的代谢比例理论(•et al ., 2003)。
在与大量的分类单元的社区,“投资组合效应”可能防止社区过程速率变化的干扰,即使类群丰富和变化不是功能冗余(Doak et al ., 1998)。投资组合效应可以发生在积极响应的一些类群平均其他类群的负面反应,导致没有净变化函数。因此,大类群的数量执行一个过程,更多的缓冲过程对环境扰动(丢弃,1995)。这种组合机制(除了功能冗余)可能导致社区相似度函数尽管微生物组成的变化。
结论
有三种方法,微生物成分可能不会对生态系统功能的重要障碍。第一,微生物群落可能难以改变。然而,我们发现的微生物组成,在大多数情况下,我们回顾了敏感二氧化碳升高,矿产受精,温度变化和C的修正案。第二,微生物成分可能有弹性,迅速回到原来的状态。文献表明,至少在几年的时间尺度,微生物组成通常还不同于安静的社区。第三,即使微生物组成变化,新的社区可能功能类似于原始。虽然目前很难测试这个假说,但最近的研究表明,许多微生物类群社区可能不是功能冗余和不同群体并不功能相似。
微生物组成的信息如何影响生态系统功能被用于改善生态系统过程的预测率扰动下吗?在前一节中给出的简单的模型我们强调两个缺少的信息。首先,需要更多的数据干扰微生物类群的反应,除了生理特征的知识。微生物类群可能不同的反应不同的干扰,而这些类群可能不对应功能组定义为生理特征。
第二,它很想知道微生物之间的关系发展史,生理特征,和响应曲线。尽管很明显,类群的系统发育关系并不完美的预测因子micro-bial生理学(阿肯巴克都和科茨,2000;Konstantinidis Tiedje, 2005),有系统信号的生理特征(如。菲勒et al ., 2007)。这些特征的遗传规模集群会指导建模者聚集微生物类群的模型。同样的,我们不知道地址之间的关系的研究发展史和微生物对干扰的反应。例如,也许微生物类群的反应特定C修正案是在非常好的系统可预测尺度(如16 s rDNA相似度> 99%),而分类单元的反应温度变化可以在更广泛的范围内聚合(如相似度> 95%)。
总之,越来越认识到微生物生态系统相关流程和巨大的进步在描述微生物的反应成分干扰,尤其是在土壤。尽管取得了这些进步,微生物生态学领域缺少强有力的预测框架来解释功能微生物成分变化的后果。需要更多的实证研究来定义
166 /史蒂文·d·埃里森和詹妮弗·b·h·她微生物功能群和他们对各种扰动的反应。更大的努力改善培养技术和评估微生物种群在控制条件下的生理反应会特别有用。一旦明确纳入模型中,这些信息可以大大提高我们的能力来预测生态系统对全球变化的反应。
确认
我们感谢德文郡布拉德利,克莱尔Horner-Devine,亚当她评价早期版本的手稿和弗朗西斯科Ayala约翰Avise,斯蒂芬·哈贝尔组织Arthur M。赛克勒讨论会“在进化之光2:生物多样性和物种灭绝。”Support to J.B.H.M. was provided by the National Science Foundation and the Gordon and Betty Moore Foundation.
继续阅读:显生宙的灭绝率下降
这篇文章有用吗?
读者的问题
-
alarico7个月前
- 回复