什么是昆虫迁徙?如何预测昆虫迁徙
一般来说,昆虫迁徙和其他运动之间的区别是人为的(Baker 1981;小费1995)。所有类型的昆虫运动的常数参数是栖息地的变化。正如Dingle(1978)所指出的那样,栖息地的改变不仅是对可能恶化的旧栖息地的逃离,而且是对新栖息地的殖民以及随之而来的更好繁殖的潜力。这一观点很早就出现了,因为Southwood(1960)没有将迁移运动与分散分开,而Andrewartha和Birch(1954)在“分散”一章下讨论了迁移。其他作者,如Rainey(1960)认为迁徙是种群的季节性迁移,而Kennedy(1961)认为它是一种持续的、明确的昆虫运动,同时抑制了阻止它的因素。Williams(1958)指出,迁徙是昆虫向一个或多或少确定的方向连续运动,而这个方向是在动物的控制下进行的。施耐德(Schneider, 1962)回顾了旧的著作,并将迁移概括为一种分散;特别是在处理有翼蚜虫的飞行时,他使用了“分散迁移”这个术语。许多作者已经认识到,这些定义仅在特定的分类群体中是可接受的概括,因为它们涉及不同群体之间的生态和动物行为学方面。
与其试图准确地定义什么是迁移,不如设置关于其组成部分和相关的迁移参数的讨论框架,从而提供一个不那么严格的定义。在昆虫迁徙中,人们可能必须首先定义一个时间成分。通常情况下,昆虫迁徙这个术语是用来描述每一个方向花费不到一年的时间,并且至少涉及两代昆虫的迁徙。地中海地区常见的一些垂直迁移,如几种昆虫的山顶迁移,是这一规律的一个例外,因为它们只涉及一代人。在这种类型的运动中,我们不能排除登上交通工具——例如,蝴蝶在大型水体中使用船只作为运输工具(Darlow 1951;Vanholder 1996)-或风的帮助(Brower 1995)。第二个因素是纯粹的空间因素,涉及昆虫移动的距离。这个距离可以从几米到几百公里不等,例如Danaus plexippus(林奈)[鳞翅目,丹奈科]和Schistocerca gregaria(福斯卡尔)[直翅目,蝗科](Tipping 1995)。Drake等人(1995)详细阐述了将昆虫运动定义为迁徙所需的要素,并在“整体概念模型”(=HCM)的背景下定义了迁徙。这个模型包含四个组成部分:[1]迁移舞台,这是发生特定迁移的垂直和水平空间; [2] the population trajectory, which involves the demographic parameters in space and time of the migrant and the receptive or the newly established populations; [3] the migration syndrome, which involves all the physiological, behavioral and morpho-anatomical traits that are associated with migrants; [4] the genetic complex, which is actually the ecology of the genes that are responsible for the traits comprising the migration syndrome. It is believed that the HCM can serve as an organizing framework for all those studies involved with insect migration. For instance, it has been stated (Mendel 2000) that the pine processionary moth Thaumetopoea pityocampa Den. & Schif. [Lepidoptera, Thaumetopoeidae] migrated in Europe from Africa by following the spread of Mediterranean pines - sensu Klaus (1989) - through land bridges that joined Europe and Africa 20 Ma BP. It has been reported that the Sahara at those times was occupied by Mediterranean-植被类型.虽然昆虫可能表现出局部迁移行为,但这种类型的进化迁移不能包括在这里给出的昆虫迁移一词中。在其他情况下,如果移动方向未知,则无法准确地识别迁移。例如,1976年英国东海岸瓢虫的大量出现被认为是来自欧洲大陆的瓢虫科(鞘翅目)的迁徙。然而,事实证明,这些掠食性甲虫只是寻找食物,并在海岸居住,因为它们被大海阻挡了,它们无法穿越(Majerus和Kearns 1989)。
在地中海国家,蝴蝶的垂直迁徙和登上山顶或多雷竞技手机版app或少是已知和可预测的事件。在这些国家,特别是雷竞技手机版app那些地形起伏丰富的国家,农民和园丁收集大量栖息在小山顶的瓢虫是一种常见的做法(Petrakis和
Legakis珀耳斯。奥林匹克广播服务公司)。所有这些活动只有一个日历基础,因为它们不是从昆虫参数的检查中推断出来的。
环境参数是最早被认为控制昆虫迁徙时间的因素之一。Kennedy(1961)指出,迁徙的沙漠蝗虫S. gregaria在种群密度较高时进入一个发育阶段,称为gregaria阶段。抑制幼年激素的释放是造成这种发育的原因。在高浓度时,幼体激素迫使昆虫进入孤期。这些变化与降雨的开始和结束相吻合;对沙漠蝗虫的生理和环境效应的耦合似乎很紧密。
几位作者指出,有些昆虫有迁移的倾向,因为它们具有运动所必需的特征。丁格尔(1996,另见1985)将这组特征称为“迁移综合征”。该综合征表现为发达的飞行肌肉组织;产卵前期较长;在新栖息地建立高繁殖力,最终有少数个体在迁徙中幸存下来;逃跑倾向;主、副飞行肌无组织分解;昆虫种群内和种群间具有较高的多态性和可塑性;相对较高的能量供应迅速建立起来,以恢复他们的迁徙。
飞行装置的多态性通常是以雄性和雌性昆虫储存的能量为代价来维持的(Zera和Denno 1977)。通常多态性有两种类型:[1]翅膀多态性和[2]飞行肌多态性。这两种类型都涉及翅膀和肌肉的数量和长度的退化。从昆虫中提取了幼体激素(JH)和蜕皮激素(ED),并对两种类型的多态性进行了分析。在最近的一项研究中,Dingle和Winchel(1997)发现soapberry bug Jadera haematoloma (Heteroptera, Rhopalidae)在翅膀长度上保持了一种复杂的多态性,涉及四种不同的变体,从大翅目(长翅)到短翅目(短翅)和小翅目(没有翅膀)。在佛罗里达群岛(主要是在基拉戈岛)的四种变种共存中,JH不仅在基因上控制变种的产生,而且在雌性的呼唤行为上发挥着重要作用,它直接影响着昆虫和一些蚜虫的迁徙飞行。有趣的是,变异的存在也受到食物的可获得性的调节(Dingle和Winchell 1997)。在新热带地区的同类昆虫Jadera aeola Dallas中,人们发现,变异是对环境线索的反应,如食物的可获得性和雨季的插入(Tanaka和Wolda 1987)。这些事实表明,在这些昆虫中,迁徙是由外部u> CD调节的
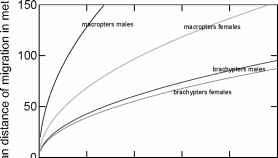
图1所示。大型和短吻鳄的平均散布距离,雄性和雌性在它们生命的前五十天。根据Socha和Zemek(2003)的公式,通过JH机制估算食物可得性等因素。在内部,根据Fairbairn(1994)调控JH的模型,JH系统可能通过加速JH的生物遗传途径和JH酯酶对JH的分解来发挥作用。
在飞行肌肉组织方面,多态更为灵活,因为飞行肌肉可以进行组织分解(Zera和Denno 1997)。对于皂莓虫也是如此(Dingle和Winchell 1997)。Andersen(1993)发现双voltine Scandinavian pond skater (Heteroptera, Gerridae)中大鳍鱼的比例一年四季都有变化。未能发现翅膀多态性与迁徙之间的相关性,是由于飞行肌肉的不同程度的组织分解掩盖了大翼龙在格里科飞行行为中的作用。如果巨肢类比短肢类有更高的飞行倾向,那么在行走昆虫中保留翅膀多态性的意义是什么?Socha和Zemek(2003)解决了这一问题,发现大翅目昆虫是Pyrrhocoris apterus (L.) (Heteroptera, Pyrrhocoridae)的一种适应性昆虫,其中[1]比短翅目昆虫具有更高的传播活性,[2]在新的区域传播和建立更好,[3]具有更高的平均速度和角速度,[4]移动距离更高(图1)。所有这些都发生在行走的运动模式中。此外,Socha et al.(1998)发现在P. apterus长距离传播之前总是有一个禁食期。通过这种方式,昆虫进入非滞育抑制繁殖,性活动非常低,这是将所有资源用于迁徙的先决条件。这些抑制,除了给昆虫时间分散之外,都发生在短臂目昆虫进食和繁殖活跃的时候。 Flightlessness of P. apterus is believed to have evolved secondarily and did not result in the loss of the physiological mechanisms that mobilizes the energy necessary for flight. Only the mode of dispersal has changed to the less costly walking that does not need the maintenance of flight muscles.
性 |
男性 |
女性 |
机翼变形 |
||
Macropter |
156.0±81.1 |
83.1±43.2 |
Brachypter |
51.8±26.9 |
47.5±24.7 |
继续阅读:昆虫垂直观察雷达VLR
这篇文章有用吗?