棕囊藻属的状态但是此属的生命周期在生态知识和既定的角色
Marie-Josephe Chretiennot-Dinet•
安妮塔·雅各布森•彼得真实性•斯图亚特·惠普尔
收到:2006年3月9日/接受:2006年5月19日/网上公布:2007年4月13日©Springer科学+商业媒体,帐面价值2007
文摘尽管许多调查,的数量和作用形态类型参与的生命周期棕囊藻属物种,但是此属仍在争论中。这部分是由于应用不同的方法如光、透射、扫描电子显微镜和流式细胞术在特定的样本。这种异质性的方法导致的不完整morpho-metric描述不同的细胞内现有一个物种根据相关标准和倍性水平的不确定观察到的每个阶段。我们审查的不同形态类型观察到在每个六棕囊藻属物种,但是此属公认。四种不同的细胞类型已被观察到。在所有六种常见的发生
绿党des系统Aquatiques ULB, CP 221大道du凯旋,1050年布鲁塞尔,比利时电子邮件:(电子邮件保护)
蔡明俊。Chretiennot-Dinet
Biologique观察站也UMR 7621实验室。阿拉戈,BP 44 66651 Banyuls-sur-Mer,法国
a·雅各布森
卑尔根大学生物学系,邮政信箱7800,挪威卑尔根5020
Skidaway海洋学研究所
佐治亚州萨凡纳10海洋科学,31411,美国
鳞状鞭毛虫生产恒星形成细丝(所有物种p . jahnii除外)(p . globosa和p . jahnii)。在三个克隆形成物种,p . globosa p pouchetii p .南极洲,三种形态类型观察:一个鞭打尺度和细丝,殖民细胞和鞭毛虫缺乏尺度和细丝。在non-colony-forming物种,p . scrobiculata和p . cordata只有鞭毛虫尺度和丝已经被观察到。怀疑在p pouchetii和p .南极洲,haploid-diploid生命周期只有证明了p . globosa。这个循环的两个主要突出特点是在殖民地布鲁姆形成和终止性是普遍存在两种类型的营养生殖。交替的生态相关性单倍体和二倍体阶段没有明确现有的生态研究的基础上。
关键词生态位•Haploid-diploid••生命周期阶段形态类型•棕囊藻属物种,但是此属•性过程
介绍
属棕囊藻属是一个全球colony-blooming但是此属物种在生物——一个重要的角色地球化学循环(Schoemann et al . 2005年)包括全球硫循环(丽丝et al . 1994年)。尽管许多调查致力于生态生理学和殖民地的角色和影响生态系统过程,属的一些主要的生物特征的知识仍然是有限的。这样的情况是棕囊藻属生命周期及其控制机制,但是此属50年Kornmann(1955)经典论文后,仍在争论中。问题分类混乱,缺乏很好的形态学描述不同的细胞类型在一个物种内,和细胞命名不足杜绝棕囊藻属生命周期。但是此属的一个完整的理解
年初以来的描述在1893年属Phaeocys-tis Lagerheim,包容的物种的数量一直是讨论的问题(例如Kornmann 1955;Kashkin 1963;帕克et al . 1971;Sournia 1988;Medlin et al . 1994年)。这主要是因为标准用来区分Phaeocys-tis物种是基于表型特征的形态测量学等殖民阶段,和/或生理生化特性(Jahnke和鲍曼1987;鲍曼et al . 1994;Vaulot et al . 1994年)。六种现在公认的基于小亚基(四)rDNA序列分析和形态学特征。这些是:p .南极洲Karsten p globosa Scherffel, p . pouchetii Hariot Lagerheim, p . jahnii Zingone, p . scrobiculata Moestrup和p . cordata Zingone et Chretiennot-Dinet (Moestrup 1979; Medlin et al. 1994; Zingone et al. 1999; Edvardsen et al. 2000; Lange et al. 2002). Colonial forms have been reported for the first four species. It is now considered that probably more than six Phaeocystis species exist (Lange et al. 2002; Medlin and Zingone this issue).
比较一个物种内的细胞类型现有使用的形态学描述标准,即。存在/缺乏身体尺度、鞭毛haptonema和恒星形成细丝,倍性水平了(例如Zingone et al . 1999;Peperzak等等。2000年)。然而,一个完整的研究的形态类型发生在一个物种失踪(兰斯洛特和卢梭2002)。我们当前的知识棕囊藻属细胞类型依赖于复合但是此属独立调查结合光(LM)、透射电镜(TEM)和扫描电镜(SEM)以及流式细胞术。这些方法提供的部分信息需要一个完整的形态类型的识别。LM有助于观察细胞的形状、大小、数量、鞭毛,和游泳活动。SEM和TEM与更高的分辨率和放大所需细胞的形态学和超微结构的细节报道,附件和细胞器。流式细胞术需要确定每个细胞类型的倍性水平。此外,至关重要的是,样品保存和固定程序完全因为这样描述程序可能导致方法论的偏见。 Use of fixatives can indeed cause cell shrinkage, loss of appendages (Peperzak et al. 2000a; Wassmann et al. 2005) or colony disintegration, releasing colonial cells into the medium, and therefore lead to possible misinterpretation (Wassmann et al. 2005). This mixed approach results in a confuse nomenclature of the various cell types, i.e. solitary flagellates and nonflagel-lates, free-living single cells, colonial flagellates, motile free-living cells, swarmers, zoids, microfla-gellates and microzoospores. These terms are often used loosely, and this can lead to misinterpretation of life cycle events.
细胞参与的数量和角色的生命周期六个棕囊藻属的物种,但是此属和是否这些是相同的在每个物种,仍在主要问题还没有解决(兰斯洛特和卢梭2002)。特别感兴趣的两个殖民地之间的识别阶段持续开花事件,以及细胞克隆形成的本质。持久性的棕囊藻属,但是此属两个殖民地之间的鞭毛虫花朵已经建议(Kornmann 1955;帕克et al . 1971;Veldhuis et al . 1986;真实性et al . 1988 b),但描述的类型的鞭毛虫从未从野外观察由于其低细胞密度和可能的混淆与其他nanoplanktonic物种。另一方面,衰老殖民地或越冬总量也被建议作为形式的p . globosa (Cadee 1991)。仍然未知因素负责生活之间的过渡阶段。不同生命周期阶段的生态意义,鞭毛虫和殖民地,最近讨论了真实性和Medlin (2003)。然而进一步调查是需要区分不同的鞭毛虫在一些物种已确定。
在本文中,我们审查和合成获得的信息领域和文化观察的不同形态类型发生在每个六棕囊藻属的物种,但是此属关注细胞形态学和倍性水平。我们还展示了未发表的数据形态的p . globosa和南极洲p细胞。生命周期内的不同形态类型的角色将根据现场和文化的观察他们的出现。最后,生活无拘束的和殖民阶段的生态相关性将讨论基于知识的生理和营养意义的各种形态类型。因为它是最著名的物种,p . globosa将被用作一个模型。
棕囊藻属物种,但是此属之间的形态类型
本节回顾了不同细胞类型报告六个棕囊藻属物种,但是此属被认为是在这里根据修订taxo-nomic地位鲍曼et al .(1994),推荐的Medlin et al。(1994)和Vaulot等等。(1994)。这特别适用于物种globosa,长期以来一直被称为pouchetii先前文献中(例如,帕克等等。1971;凯塞1970;Admiraal和Venekamp 1986;Veldhuis等等。1991;戴维森和马尔尚1992)。不同细胞类型的季节性分布在自然环境中也会被认为是为了评估他们在生命周期中的作用。
p . globosa的形态类型
仔细分析的文献出版以来的第一次描述p globosa细胞Scherffel(1899, 1900)表明,四种形态类型存在:二倍体殖民细胞,二倍体鞭毛虫,两种类型的单倍体鞭毛虫。
殖民时期的细胞
殖民细胞有2 - 4顶叶叶绿体剥夺身体尺度,haptonema、鞭毛和嵌入在一个粘的矩阵(Scherffel 1899;Kornmann 1955)。他们拥有鞭毛极两个短的附属物,是目前未知的作用和性质(图1;卢梭等人提交)。报告为殖民生活细胞大小范围4.5 | -8.0米(Kornmann 1955)和5.8 | -10.4米(卢梭等人提交),而他们是5.68.3 | m (Peperzak et al . 2000年)和4.6 | -7.8米(卢梭等人提交)Lugol和Lugol-glutaraldehyde固定细胞。这些细胞是二倍体(Cariou et al . 1994;Vaulot et al . 1994年)。他们是均匀分布在殖民地15 - 20 |米下一层薄薄的皮肤,并与稀弱互联凝胶(Kornmann 1955;范Rijs-sel et al . 1997;哈姆et al . 1999年)。 The colony skin is strong and semipermeable with pore size 1.04.4 nm in diameter, and presents plastic and elastic properties (Hamm et al. 1999). Diameter of colonies typically ranges from 10 |m to 8-9 mm (Kornmann 1955; Jahnke and Baumann 1987; Rousseau et al. 1990), but may occasionally reach 20-30 mm (Kayser 1970; Gieskes and Kraay 1975; Chen et al. 2002). Colonies are originally spherical but deviate into nonspherical shapes when growing larger or when subjected to hydrodynam-ical stress (Kornmann 1955; Bätje and Michaelis 1986; Rousseau et al. 1994).
Non-motile自由细胞从殖民起源也被报道在p globosa文化。这些细胞形态相似的殖民细胞:同样的尺寸范围,缺乏鞭毛,haptonema (Kornmann 1955;卢梭et al . 1990;1994;Peperzak 1993;Peperzak et al . 2000;Dutz和人类研究2006),身体尺度(Peperzak et al . 2000年),形如细丝和星星(Peperzak et al . 2000;Dutz和人类研究2006)和被认为可以有相同的倍性水平,即。二倍性(Peperzak et al . 2000年)。在此基础上,non-motile生活无拘束的和殖民细胞应该不再被视为不同的形态类型(Peperzak et al . 2000;Dutz和人类研究2006)。
哈普罗id鞭毛虫
鞭打的精细结构阶段p globosa首次被描述为从TEM p . pouchetii帕克et al . (1971)。这些细胞已经被报告为蜂群(Scherffel 1900), micro-zoospores (Kornmann 1955),小型和大型zoids(帕克et al . 1971), microflagellates
图1的SEM照片p globosa细胞中观察到比利时的沿海水域。与恒星和小型Xagellates Wlaments但没有尺度观察(一)pre-bloom期间2月;酒吧= 3 ^ m和(B)在可能仍然嵌入到殖民地粘液的最后殖民阶段;酒吧= 2 ^ m;殖民细胞典型的短附观察(C)的早期阶段内开花Chaetoceros刚毛;酒吧= 4 ^ m和(D)的最后开花前形成单倍体Xagellates;酒吧= 2.4 m ^。H: hapto-nema;Fl: Xagella;S:明星; F: Wlament; M: mucus; A: short appendage; Ch: Chaetoceros cell"
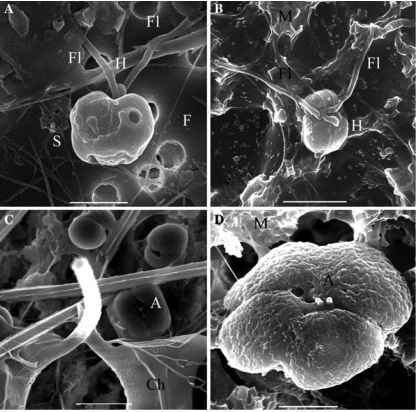
(Veldhuis et al . 1986年)和微观,mesoflagel-lates (Peperzak et al . 2000年)。他们都是圆形的,而且比殖民细胞小,直径3 - 5 m ^ (Kornmann 1955);3 - 6 m ^(帕克et al . 1971年)和3.6 - -5.8 m ^(卢梭等人提交)LM下活细胞观察。小尺寸一直被用于区分他们从其他p globosa细胞类型(例如Veldhuis et al . 1986年)之前Vaulot et al。(1994),使用流式细胞仪,展示他们的单倍体阶段p . globosa。
这些鞭毛虫能够快速营养生殖(Kornmann 1955;帕克et al . 1971;卢梭et al . 1994;Vaulot et al . 1994年)和游泳很积极(Kornmann 1955;帕克et al . 1971年)。他们拥有两个等于heterody-namic鞭毛,10 - 15 ^米长度和短haptonema(3 - 4 ^米)远端肿胀。他们呈现的是一个前抑郁和两个金叶绿体(帕克et al . 1971年)。胞体是由两种类型的有机尺度显示辐射山脊的模式,可见双方(帕克et al . 1971;
Peperzak等等。2000年)。两种类型的单倍体鞭毛虫被帕克杰出et al . (1971)。一种类型,3 - 6 ^米大小,拥有两个表面,明亮的囊泡位于人体表面(帕克et al . 1971年)。这些囊泡释放细丝,20 m ^, ^ 0.05米直径,并由alpha-chitin (Chretiennot-Dinet et al . 1997年),形成一个高度特征五角星星(帕克et al . 1971年)。这些细丝的功能尚不清楚。被假定他们可以作为附件的锚固体结构前殖民地开始(Chretiennot-Dinet 1999),或者有一个角色在防御食草动物(Peperzak et al . 2000;Dutz和人类研究2006)。单倍体鞭毛虫,3 - 5 ^米大小,缺乏囊泡和细丝(帕克et al . 1971;Peperzak et al . 2000年)。 Such a difference in size and ability to produce filaments has also been observed in field samples for cells described as zoids (Manton and Leadbeater 1974 cited in Peperzak et al. 2000a) or meso- and micro flagellates (Peperzak et al. 2000a).
这些鞭毛虫在衰老文化,不同大小的球形殖民地内游泳,与殖民地消失(Kornmann 1955)。在自然环境和昌盛,大量的这些鞭毛虫被多次报道,在p . globosa殖民地花朵的衰落,在球形殖民地(Scherffel 1899, 1900;Cadee 1991;Peperzak等等。1998、2000),或释放到介质离开鬼殖民地或独立生存的鞭毛虫(琼斯和哈克1963;Admiraal和Venekamp 1986;Veldhuis et al . 1986;真实性et al . 1988 b;Escaravage et al . 1995年)。这些小型鞭毛虫观察总是与几丁质的丝和星星的存在有关,p的单倍体阶段特有的特征globosa Scherffel (Vaulot et al . 1994; Zingone et al. 1999). Released from colonies at the end of the bloom, these flagellates were recorded at different periods of the year in the water column of the Channel and the Southern Bight of the North Sea, at cell densities varying from 80 x 103 cells L-1 to 220 x 103 cells L-1 (Fig. 1; Rousseau and Chretiennot-Dinet, unpublished data). They apparently represent the life stage persisting between two blooms of colonial cells, as suggested by Kornmann (1955) and Parke et al. (1971).
二倍体鞭毛虫
p . globosa第三形态类型是相同的尺寸范围的鞭毛虫殖民细胞观察生活时,即4.5 - -8.0 |米(Kornmann 1955), 6 - 7 | m (Peperzak et al . 2000年)和6.1 | -9.3米(卢梭等人提交)包含两个鞭毛和一个haptonema但缺乏规模,丝和星星(卢梭等人提交)。这鞭毛虫被Kornmann报告为无性繁殖的游动孢子(1955),作为一个大型鞭毛虫Cariou et al。(1994)和卢梭et al .(1994),作为一个macroflagellate Peperzak et al。(2000)。它通常出现在文化在24 h当殖民细胞释放机械从殖民地(Kornmann 1955;卢梭et al . 1990;Cariou et al . 1994年)。这鞭毛虫是二倍体没有发现倍性变化在殖民细胞转变成鞭毛虫(Cariou et al . 1994;卢梭et al . 1994年)。
这些鞭毛虫能够迅速形成新的殖民地后一天内粘附到表面(Kornmann 1955;凯塞1970;Cariou et al . 1994;卢梭et al . 1994年)。无生命的粒子(卢梭et al . 1994),文化的血管壁上(凯塞1970;Cariou等等。1994年),和硅藻(weisse等等。1986;Boalch 1987;卢梭等等。 1994) have been observed as adhesion sites. This property of attachment to surfaces, specific to this life stage, led to the assumption that a benthic stage, acting as an overwintering form, exists in the natural environment (Kayser 1970). It is not clear if this flagellate is able to mitotically divide. Its rapid transformation into a colony suggests it is short-lived (Kornmann 1955; Rousseau et al. 1994), but vegetative multiplication was also reported by Kayser (1970). The short lifespan of this morphotype could well explain why it is observed only occasionally in the field (Kornmann 1955; Peperzak et al. 2000a; Rousseau et al. submitted).
这样的鞭毛虫偶尔观察到内部殖民地文化的Kornmann(1955)认为他们为不同的细胞类型,macrozoospores,在自然环境中(Peperzak et al . 2000年)。
p . pouchetii的形态类型
目前,两个p pouchetii细胞类型已经证实基于EM观察和仪分析:一个二倍体殖民细胞和一个二倍体鞭毛虫(雅各布森2002)。然而,其他报告表明,一个更大的鞭毛虫p . pouchetii内可能存在生命周期(Sukhanova和燧石2001;Wassmann et al . 2005年)。
殖民时期的细胞
二倍体殖民细胞尺寸范围的57 | m,前纵槽和被剥夺的丝状的附属物和规模覆盖物(雅各布森2002)。积极发展殖民细胞群体的叶通常分布在云计算殖民地(Jahnke和鲍曼1987;Gunkel 1988;鲍曼et al . 1994;卢梭et al . 1994;雅各布森2002)。棕囊藻属pouchetii但是此属殖民地,球形到0.1毫米(卢梭et al . 1994年),有一个最大尺寸1.5 - 2毫米(Jahnke和鲍曼1987)。他们的特点是一个微妙的粘液,容易破坏相比,p . globosa固体粘液(van Rijssel et al . 1997;雅各布森2002;Wassmann et al . 2005年)。Non-motile自由细胞形态类似于殖民时期的细胞,即。,剥夺鞭毛haptonema丝状附属物,尺度,可以找到一起殖民地阶段由于殖民地中断(Eilert-sen 1989;雅各布森2002;Wassmann et al . 2005年)。
鞭毛虫
详细描述了一个filament-bearing鞭毛虫雅各布森et al。(1996)和雅各布森(2002)LM和TEM观察的基础上。鞭打,源于殖民地时带进文化,是圆的,平均直径5我当生活。它有两个金壁叶绿体,两个hetero-dynamic同样长(11 |米)鞭毛,和一个短non-coiling haptonema(| 1.5米)。胞体是由两种类型的尺度,与辐射山脊两个表面上可见。丝缠绕在一个或两个表面的囊泡,并形成five-ray恒星时解除。p . globosa filament-producing细胞的相反,这鞭毛虫是假定为二倍体(雅各布森2002)。观察该鞭毛虫在冬天,之前增加丰富的殖民开发,被认为是殖民的前兆阶段(雅各布森2002;雅各布森和Veldhuis 2005)。这样鞭毛虫观察内部殖民地在春天的花朵,在随后发布的水柱(雅各布森2002)。
LM观察的样品,两种类型的鞭毛虫被报道在p pouchetii生命周期基于尺寸标准,即。,大(6 |米;Sukhanova和燧石2001;Wassmann et al . 2005年)和小的心形(34 |米;Wassmann et al . 2005年)细胞。大型鞭毛虫跟殖民地和丰富,有时是占主导地位的殖民细胞的细胞密度(Sukhanova和燧石2001;Wassmann et al . 2005年)。发现小鞭毛虫独立生存的(Wassmann et al . 2005年)或在消失前的腐烂的殖民地(Lagerheim 1896;Wassmann等等。2005)。他们很可能对应于描述的鞭毛虫雅各布森(2002),与大小差异解释为细胞收缩由于使用Lugol-glutaraldehyde样本固定剂。 Interestingly, small flagellate formation and their further release into the water column have been shown to originate from colonies assuming a spherical shape (Gunkel 1988; Whipple et al. this issue).
形态类型的南极洲
三种形态类型曾被观察到在南极洲p:殖民细胞和两种类型的鞭毛虫。两倍性水平已经被记录在案。
殖民时期的细胞
殖民p .南极洲没有鞭毛,细胞没有haptonema,没有鳞片,没有小泡或恒星形成细丝(戴维森1985)。这个细胞有一个前抑郁和两个短的附属物globosa殖民页所观察到的类似细胞(图2;Chretiennot-Dinet和卢梭,未发表)。殖民细胞的尺寸范围报道p南极洲是相当大的,根据保护和固定程序。尺寸范围包括:10我活细胞(Moestrup和拉森1992)和3.2 | -7.9米(Mathot等等。2000),4.7 - -5.6 |米(Vaulot等等。1994年),4 - 6 |米(本研究)Lugol-gluraldehyde固定细胞。殖民的细胞p南极洲,认为是dip-loid (Vaulot et al . 1994),沿周边均匀分布的殖民地特点是一个坚实的粘液。p .南极殖民地通常是球形但可以包括许多变种的最大尺寸9毫米(鲍曼et al . 1994;军火商和汤姆森1994)。
鞭毛虫
一个p .南极鞭毛虫熊尺度和造成丝和星星;另一种是没有尺度,丝和星星。详细描述了scale-bearing鞭毛虫戴维森(1985):它有一个前抑郁,两个chlorop-lasts中央大蛋白核,和熊两个鞭毛和haptonema。细胞大小范围在3.5 m和7 | |米时固定
图2 p .南极洲的SEM照片(1871年应变CCMP):(一)殖民细胞有四个叶绿体和两个短附典型的殖民鞭毛杆细胞;酒吧= 2点:(B)鞭毛虫共病的殖民地(二倍体)剥夺尺度和细丝;酒吧= 6点。H: haptonema;Fl:鞭毛;答:短肢
图2 p .南极洲的SEM照片(1871年应变CCMP):(一)殖民细胞有四个叶绿体和两个短附典型的殖民鞭毛杆细胞;酒吧= 2点:(B)鞭毛虫共病的殖民地(二倍体)剥夺尺度和细丝;酒吧= 6点。H: haptonema;Fl:鞭毛;答:短肢
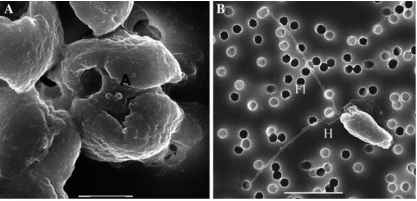
戊二醛和TEM观察到。同样两个鞭毛长(6 - 10点)。haptonema呈现球状的小费,是1.5下午2点。人体细胞是由两种类型的尺度模式的辐射山脊两侧可见。线型材料定期安排内部循环posterio-lateral囊泡。部署时,线程的长度25点,形成一个五边形的明星。
在自然环境中,这种scale-bearing鞭毛虫增加数量在殖民的开始开花,开花期间与衰落;他们存在于大量开花后(戴维森和马尔尚1992 b)。类似鞭毛虫,直径5.2 pm与戊二醛固定时,两种鞭毛的14.3点的长度,和丝状附属物40点的长度形成一个五边形的明星,在被发现冰边缘威德尔海在南国的夏天(1983年巴克和驻军)。鞭毛虫线型材料观察伯兰斯菲尔德海峡地区post-bloom期间(1995年康和李)。另一方面,增加大量的小型独立生存的鞭毛虫(2.4 - -5.5 pm与戊二醛固定时)在月初罗斯海南国春集落形成之前发生(Mathot et al . 2000;史密斯et al . 2003年)。虽然没有提供良好的形态描述,这些小鞭毛虫可能对应于描述的鞭毛虫巴克和驻军戴维森(1983)和(1985),大小差异造成细胞收缩和准备显微观察。他们的相对丰度下降到最低,而殖民细胞主要在春季晚些时候他们再次观察到内部殖民地(史密斯et al . 2003年)或自由细胞(推杆et al . 1994年)结束的时候开花。
第二个p .南极鞭毛虫殖民细胞类型有相同的大小,从6.5点(加里森和汤姆森1993)到7.5点弗里克塞尔(1989)。这个鞭毛虫熊两个鞭毛和haptonema但缺乏规模,丝和明星(图2、驻军和汤姆森1993;军火商和汤姆森1994;Chretiennot-Dinet和卢梭,未发表)。这鞭毛虫成立内部和释放球形或细长的殖民地冰边缘在南国之春(弗里克塞尔1989;加里森和汤姆森1993;军火商和汤姆森1994)。一些5 - 6 h形成后,发现连接到大型硅藻的刺和过程(加里森和Thom-sen 1993;军火商和汤姆森1994),随后形成新的殖民地弗里克塞尔(1989)。
p . jahnii的形态类型
两种细胞类型描述了p . jahnii:殖民细胞和鞭毛虫(Zingone et al . 1999年)。
殖民时期的细胞
棕囊藻属jahnii但是此属殖民地细胞具有两个短附典型的p . globosa和p .南极殖民细胞,有大小6 - 8.5点当生活和不规则地分布在松散,不规则的殖民地。Non-motile相同大小的自由细胞也被报道(Zingone et al . 1999年)。
表1不同形态类型报告六个棕囊藻属物种公认。但是此属
测量或假定的倍性水平——而不是观察到
鞭毛虫鞭毛虫鞭毛虫没有鳞片,殖民鳞片,鳞片线程和明星一样大小的细胞线程和明星范围作为殖民细胞
p . globosa p pouchetii南极洲p p . jahnii p cordata scrobiculata页
继续阅读:X
这篇文章有用吗?