维护内源性代谢衰变裂解和死亡
屈服值在前面的部分是那些结果当所有能源获得的生物质合成传经送宝。合成并不是唯一的能源要求微生物,然而。他们还必须有能量进行维护。”
细胞过程,无论是机械或化学,需要能量的表现,除非供应可用这些基本过程将停止,细胞会变得混乱和死亡。机械过程包括运动性、渗透调节、分子运输、维护离子梯度,而在一些真核生物中,胞质环流。虽然可能认为能动性可以舍弃某些微生物,这种观点并不适用于所有因为一些需要能动性去寻找食物。在所有细胞渗透调节非常重要,即使是那些受严格保护细胞壁,和泵机制,如收缩液泡,存在于细胞来抵消渗透压泵水的正常趋势。细胞膜渗透到许多小分子,如氨基酸,因为高浓度的这些倾向于扩散到细胞内的介质。主动运输机制运作这样的分子进入细胞浓度梯度。类似的自然是维护一个离子梯度的必要性穿过细胞膜,而质子动力密切相关负责ATP合成。维护这个梯度被认为是维护的主要消费能源。71最后,胞质环流和材料内部的运动Eu-carya通常所需的正常运作。他们也需要能量。
化学因素也有助于维护能源需求。微生物细胞代表化学组织和其内部的许多组件有更高的自由能比原来他们形成的化合物。一般来说,因为这个组织,能量必须在正常的倾向可以抵消障碍,即。、克服熵。化学过程导致的能源要求维护弧那些参与resyn-thesis等结构的细胞壁,鞭毛,细胞膜和异化的装置。例如,一所“建议能源的再合成蛋白质和核酸是一个重要的部分的维护能源要求大肠杆菌。
一个主要的争论点有关微生物文学对维护能源需求的影响文化的速度在增长。早期调查”表明,需要维护能源独立的增长率,但最近的研究表明相反。1不过,工程师们通常认为维护能源需要独立的增长率在污水处理和生化操作中采用的方法,将这本书。
给定的存在需要维护的能量,它可以使用能源供应什么?这个问题的答案取决于微生物的生长条件。如果一个外部(外生)能源供应是可用的,它的一部分将用于满足维护的能源需求,其余部分将用于合成。能源供应的速度下降,越来越少的将用于新增长,因此网络,或观察到的,产量将下降。当关键是到达率的能源供应平衡的能量的速度必须用于维护,没有净增长将会发生,因为所有可用的能量将用于维持现状。如果进一步减少能源供应,供应率之间的差异和维护能源需求将被满足的降解细胞内的可用的能源,即。内源性代谢。这将导致减少的质量文化。最后,如果没有外生能源可用,所有的维护由内源性代谢必须满足能源需求。当达到指向所有内生储备已经筋疲力尽,细胞恶化和死亡,或进入休眠状态。
材料作为基质的性质为内源性代谢取决于微生物的物种和文化生长的条件。例如,当大肠杆菌生长迅速在glucose-mineral盐介质储存糖原。1“如果这些细胞电弧然后放在一个环境缺乏外源底物,他们将利用糖原作为内生能源。氨基酸和蛋白质显示小净分解代谢,直到糖原。胰蛋白胨培养基中生长时,另一方面,大肠杆菌积累小糖原。因此,内源性代谢利用含氮化合物。其他生物仍然使用其他化合物,包括核糖核酸(RNA)和脂质聚(3-hvdroxybutyrate (PHB)。
一个问题困扰了微生物学家担忧时能量流的路线外源性底物可以供应充足的维护能源需求的文化。内源性代谢是否继续在这种情况下,这样的一部分能量释放基质的降解是用来resupplv国内能源储备被内源性代谢降解吗?,或者是内源性代谢停止,以便释放的能量直接从外源性底物的降解为维护功能?的证据仍不确凿。实际上,虽然这些问题是基本的科学意义,他们liltle轴承工程师使用的宏观能源平衡数学模型生化操作。事实上,当我们看到3.3.2节。一些模型避免整个问题通过引入细胞lvsis和再生的概念。
生物量的数量实际上形成单位基质用于生化操作,称为观察产量(Y„K)。总是小于y的一个原因是能量需要维护。必须消耗更多的能量,出于维护目的,这个少用于合成和较小的生物量的数量单位基质降解形成的。其他f演员也有助于区别,然而。例如,考虑捕食的影响。在一个复杂的微生物群落,如发现在活性污泥法、原生动物和其他真核生物捕食细菌,减少生物量形成的净额。为了说明捕食的影响。假设Y的值卤细菌生长在葡萄糖是()。(•>()毫克每毫克的细菌生物量鳕鱼形成葡萄糖鳕鱼。因此,如果100 mg / L的葡萄糖鳕鱼。60毫克即细菌生物量鳕鱼的结果。现在,假设Y的值为原生动物捕食细菌是0.70毫克每国家行业集团公司形成的原生动物生物量鳕鱼细菌生物量鳕鱼。如果产生的原生动物消耗所有的细菌葡萄糖,结果将会是42 mg / L(原生动物生物量。结果,如果我们只观察到的净生物量形成,没有区别,这是什么,我们会得出这样的结论:42 mg / L的生物质能鳕鱼造成的破坏100 mg / L的葡萄糖鳕鱼。因此,我们可以得出这样的结论:观察收益率为0.42。小于真实的增长产生细菌生长在葡萄糖。 Macroscopically, it is impossible to distinguish between the v arious factors acting to make the observed yield less than the true growth yield. Consequently, environmental engineers lump fliem together under the term "microbial decay," which is the most common way they have modeled their effect in biochemical operations."'
另一个进程导致的损失生物质在生化操作细胞溶菌作用。1细菌的生长需要协调的细胞壁的生物合成和降解材料允许细胞扩张和分裂。负责的细胞壁水解的酶称为自溶素和他们的活动通常是在严格监管下允许他们一致行动与生物合成的酶在细胞分裂。损失的规定,然而,会导致破裂的细胞壁(溶解)和死亡的生物体。当细胞壁破裂,胞浆和其他内部成分的释放中,他们成为其他生物生长基质的文化。此外,细胞壁和细胞膜,以及其他结构单元,开始受到水解酶在中,增溶的他们,使他们可以作为基质。只有最复杂的单位仍然ccll碎片,这是可溶性这么慢,它似乎是耐火材料在大多数生化操作。4 ^ 4的参数如何溶解导致的损失生物量与捕食相关类似,上面说明。收益率表现出由细菌生长在可溶性产品发布的裂解是相同大小的收益率与其他生物基质上的生长。因此,如果100 mg / L的生物细胞溶解,仅50 - 60 mg / L的生物将会从re-growth裂解产品。因此,溶解和再生的净效果是减少系统中生物量。 In general, starvation itself does not initiate lysis, although the events that trigger it are not yet clear. Nevertheless, engineers seeking to model the decline in observed yield associated with situations in which the microbial community is growing slowly have focused on cell lysis as the primary mechanism.12 2"
最后的事件影响的活性生物生化操作是死亡。传统上,死细胞已经被定义为一个失去了分裂的能力在琼脂板”2和基于这个定义的研究表明大部分微生物的缓慢增长的文化是不能存活的,或死亡。“1””4此外,总结维德尔和詹金斯毫升大量研究使用间接证据涉及基质去除率和酶活性的比较得出结论,大部分的美国职业足球大联盟在废水处理系统是不活跃的。然而,后来的一项研究中,4“41使用更复杂的技术来识别死亡细菌,建议分数很低的细胞在低增长率实际上是死了。相反,许多人只是nonculturable标准技术,尽管他们仍然活着。此外,最近work4”表明,死细胞不长时间保持不变,而是溶解,导致基质和生物残骸,正如上面所讨论的。生物质垃圾行为的存在使可行的微生物的质量低于系统中悬浮物的质量。尽管这本书的前任使用model1”明确地考虑细胞死亡,现在看来,直接考虑的现象是没有依据的。4“41相反,事实上只有一个部分的mls在生物废水处理系统实际上是可行的生物量的积累可以归因于生物量碎片而不是死细胞的存在。
总之,由于几种机制,生化反应堆展览两个重要特征:(1)观察到的收益率小于真实增长产量和(2)活跃,可行的细菌只占一小部分的“生物”。One simplified conceptualization of the events leading to these characteristics is that bacteria are continually undergoing death and lysis, releasing organic matter to the environment in which they are growing. Part of that organic matter is degraded very, very slowly, making it appear to be resistant to biodégradation and causing it to accumulate as biomass debris. As a consequence, only a portion of the "biomass" is actually viable cells. The remainder of the released organic matter is used by the bacteria as a food source, resulting in new biomass synthesis. However, because the true growth yield is always less than one, the amount of new biomass produced is less than the amount destroyed by lysis, thereby making the observed yield for the overall process less than the true growth yield on the original substrate alone.
2.4.3可溶性微生物产物的形成
废水中的可溶性有机物的微生物来源的生物反应器和由微生物降解有机基质影响的生物反应器。这一现象的主要证据来自实验中单可溶性基质的成分被喂以微生物培养物和由此产生的有机化合物废水的研究影响底物的存在。“大量的废水有机物并非原始基质和较高的分子量,这表明它是微生物的起源。这些可溶性微生物产物被认为源于两个过程,一个生长-,另non-growth-associated。生长-产品形成结果直接从生物量增长和底物的利用率。因此,它是通过另一个产量因素耦合到这些事件,微生物产品产量、YMl。,一个单位的生物降解底物生产YM1结果,单位的产品。伊夫圣罗兰的值。对各种有机化合物被发现小于0.1。“Non-growth-associated产品形成与衰变和裂解和导致biomass-associated产品。他们被认为来自释放可溶性细胞成分通过溶解和颗粒细胞成分的溶解。 Although little is known about the characteristics of these two types of soluble microbial products, they are thought to be biodegradable, although some at a very low rate. Compared to other aspects of biochemical operations, little research has been done on the production and fate of soluble microbial products and few researchers have attempted to model the contribution of such products to the organic matter discharged from wastewater treatment systems."'Ml Nevertheless, an awareness of their existence is necessary for an accurate understanding of the response of biochemical operations.
2.4.4溶解的颗粒和高分子量有机物
细菌只能占用和降低低分子量的可溶性有机物。所有其他有机材料必须受到细胞外酶释放低分子量化合物,可以跨细胞膜转运。许多有机聚合物,特别是微生物来源,如细胞壁成分,蛋白质,核酸,是由几个重复的子连接债券可以被水解。因此,微生物的过程将颗粒和高分子量的可溶性有机化合物分解成子单元通常称为水解,尽管一些涉及的反应可能会更复杂。
水解反应扮演两个重要角色为污水处理生化反应堆。首先,他们负责细胞成分的溶解释放细胞溶菌作用的结果,防止他们系统中累积。因为细胞溶菌作用发生在所有微生物系统,甚至水解反应是重要的生物反应器只接收可溶性底物。第二,许多生化操作接收微粒有机材料,在这种情况下水解带来所需的生物降解是至关重要的。尽管其生化功能的操作中的中心地位,相对很少有研究试图站下水解的动力学机制。“不过1:1,这具有重要影响的结果必须考虑生化操作和一个完整的理解的功能。
2.4.5加氨
氨化的名字是氨氮的释放氨基酸和其他含氮有机化合物进行生物降解。这是一个正常的生物降解过程的结果,在氨基酸组电弧解放和分泌细胞氨。氨化率将取决于包含基质氮利用率的速率和碳氮比的衬底。加氨氮控制污水处理过程中是非常重要的,因为有机氮不受氧化硝化细菌。他们只能后硝酸氧化氮转化成氨和释放介质。
2.4.6磷吸收和释放
如果悬浮生长生物反应器系统配置为两个系列第一区厌氧区和第二个有氧,有关报道,具有一种特殊的代谢能力不属常见细菌,会扩散并存储大量的无机磷酸盐作为多磷酸盐,从而允许通过生物质消耗量从废水除磷。尽管有关报道中经常出现在大量完全有氧暂停增长文化,他们只开发能够存储大量的磷酸盐受到交变时无氧和有氧条件被回收之间的两个区域。7这从他们独特的碳储存能力为代价的磷酸盐在厌氧条件下磷酸和存储的碳在有氧条件下。两个场景假设来解释有关的功能。是独立开发的Comeau et al。”和Wentzel et al ., K”而另一个是由阿伦et al。前者被称为Comeau-Wentzel模型而后者叫做米诺模型。K1两个模型之间的差异在有关报道中代谢多样性的结果,因为还不知道这模型是更普遍适用的,都将呈现。
Comeau-Wentzel模型。我们将首先考虑事件发生厌氧区。因为发生在下水道的发酵,大部分可溶性有机物废水在国内醋酸和其他形式的短链脂肪酸。此外,废水进入厌氧生物反应器时,多余的脂肪酸是由发酵反应由兼性异养生物。如图2.5所示,醋酸由被动扩散穿越细胞膜(如未离解的醋酸),但是一旦进入,是激活acetyl-CoAby耦合ATP水解,产生ADP。虽然没有显示在图中,ATP还用于维持质子动力传输已经失去的质子与未离解的醋酸。细胞响应减少通过刺激ATP再合成ATP / ADP比率从存储多磷酸盐(Poly-P„)。乙酰辅酶A的一部分通过三羧酸循环代谢提供减少所需功率(NADH + H)
外面在外面里面
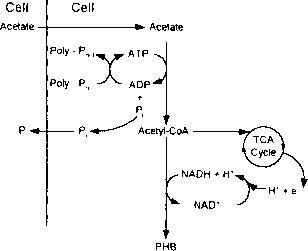
一个厌氧b .有氧
图2.5原理图描绘Comeau-Went / el模型卤无机磷酸盐的吸收和释放有关报道:a .厌氧条件;b .有氧条件。(改编自走/ el et al.sl)
PHB的合成。乙酰辅酶a的其余部分转化成的PHB,约有90%的醋酸碳被保存在存储聚合物。没有多磷酸盐的存在为ATP提供能量再合成。醋酸醋酸会在细胞中堆积,交通将会停止,会发生和PHB没有形成。多磷酸盐形成ATP水解的增加细胞内无机磷酸盐的浓度,P„释放到本体溶液,随着阳离子(图中未显示)保持电荷平衡。
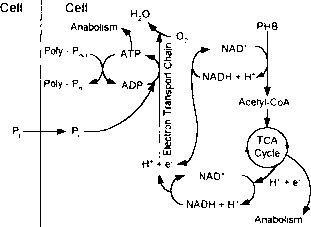
当废水和相关的生物进入有氧区,在可溶性有机物废水低,但有关包含大量的PHB储备。此外,废水富含无机磷酸,而有关多磷酸盐水平较低。因为他们在有氧区氧作为电子受体,有关执行正常增长的有氧代谢利用存储的PHB作为他们的碳和能量来源,通过电子传递磷酸化生成ATP,如图2.5所示。此外,随着ATP-ADP比率的增加,刺激多磷酸盐的合成,从而去除磷酸盐和相关的阳离子(未显示)解决方案和再生细胞存储多磷酸盐。因为提供的大量的能量存储的PHB的有氧代谢,有关报道能够吸收所有的磷酸磷酸在厌氧区+发布最初出现在废水。
厌氧和之间的不断循环有氧区给出了有关竞争优势在普通的异养细菌,因为没有能力制作和使用多磷酸盐,普通的异养生物不能占用有机质的厌氧区。应该注意的是,虽然大多数系统去除磷酸盐通过使用有关使用有氧区再生的存储多磷酸盐,另一些有关可以使用硝酸盐和亚硝酸盐作为电子受体,允许缺氧条件下要使用。
米诺模型。米诺模型,如图2.6所示。Comeau-Wentzel模型非常相似,主要区别是糖原的作用,碳水化合物储存聚合物。在这种情况下,在厌氧区所需的还原能力合成乙酰辅酶a的PHB的来自葡萄糖释放糖原的代谢。葡萄糖氧化通过Entner-Doudoroff丙酮酸(ED)或Embden-Meyerhof-Parnas (EMP)通路,PAO的类型不同,从而提供一些所需的ATP乙酸转化为乙酰辅酶a和PHB合成所需的还原能力。反过来,丙酮酸氧化]}’脱羧乙酰辅酶a和二氧化碳,释放的电子也被用于PHB的合成。因此,所有的醋酸了存储的PHB,从糖原是碳的一部分。在有氧区,PHB Comeau-Wentzel模型分解为磷酸为生物合成提供以及吸收和存储多磷酸盐。此外,然而,PHB还用于补充糖原存储。
2.4.7概述
图描绘的整体和事件的发生好氧生物反应器获得可溶性底物是如图2.7所示。细菌消耗衬底(Ssl)和生长,从而导致更多的细菌、底物消耗和生物量增长之间的关系是由真正的增长收益,y
细胞外
细胞内
继续阅读:化学计量学和广义反应速率
这篇文章有用吗?