分类研究
我们检查了大约200 Pleisocene和全新世沉积物样品,采集来自29个钻孔的深度0.5 -47,以及埋藏土壤和cryopedolith-located化石啮齿动物洞穴的材料。ice-complex沉积物,可行的原生动物被发现在125年25样品(样品检查总数的20%)。发生的可行的原生动物埋在土壤中相当高(80%;14个样本)和化石洞穴(100%;12样本)(表8.1)。
protozoological分析的样本显示标本主要原生动物macrotaxons:裸阿米巴原虫,异养鞭毛虫,纤毛虫和太阳虫目(表8.2,图8.3)。
12文化cyst-forming古代纤毛虫物种得到样品的冻土沉积物,埋土壤和洞穴。他们表示,在他们的主要部分,polyzonal物种的标本。除了十株标本分类群Colpodea——肾形虫属steinii, c . inflata c aff.粗,c . aff. augustini,肾形虫属sp, Platyophrya aff. vorax-the纤毛虫钟形虫sp (Oligohimenophorea)和Oxytricha sp。(Spirotrichea)是孤立的。
原来可行的变形生物的发生(49%),永冻层沉积物高于纤毛虫(9%)。裸体变形虫被发现在永冻层沉积物的样本和埋土。我们确定了分裂的标本(Leptomyxida Acanthamoebidae)和heterolobose (Vahlkampfiidae)阿米巴原虫。纯文化古老的裸体阿米巴原虫,两个Leptomyxida和八Acanthamoebidae菌株,得到在实验室设置。acanthamoeba,像colpodean纤毛虫,分布在全球范围内(1988页;Foissner 1993)。
安葬洞穴的样本,我们发现27个物种和形式的het-erotrophic鞭毛虫从十分类组和鞭毛虫incertae基准(Shatilovich et al . 2008年)。古代鞭打动物的分类分析显示,变形鞭毛虫(Cercomonadida Apusomonadidae)和stramenopiles(金藻纲)是最大量和不同组织(图8.4)。
网站 |
位置 |
嗯,没有。 |
深度(米) |
年龄 |
《创世纪》 |
岩性 |
1 |
拉普捷夫海海岸Bykovsky半岛 |
1/01 |
0.40 - -0.56 |
现代土壤 |
||
1 |
QIII |
晚更新世沉积物的冰复杂 |
泥炭 |
|||
1.95 - -2.05 |
砂壤土和泥炭 |
|||||
2.16 |
||||||
2.8 |
||||||
7 |
||||||
19 |
砂壤土 |
|||||
2/01 |
1.0 - -1.1 |
QIV |
沉积物的全新世唉 |
砂壤土和泥炭 |
||
2 |
||||||
2.25 - -2.33 |
||||||
2.4 |
||||||
2.5 |
||||||
7/03 |
4 |
QIII |
晚更新世沉积物的冰复杂 |
泥炭 |
||
4.95 - -5.05 |
砂壤土和泥炭 |
|||||
6.9 |
泥炭 |
|||||
12/03 |
3.5 |
QIV |
的全新世沉积物唉 |
砂泥炭的夹杂物 |
||
4 |
||||||
2 |
Yana-Indigirka低地,色度湾口 |
2/04 |
0.71 |
现代土壤 |
||
1.15 |
QIII |
晚更新世沉积物的冰复杂 |
壤土 |
|||
3 |
Indigirka-Kolyma低地, Khomus-Yuryakh河 |
3/05 |
0.7 |
现代土壤 |
||
4.2 |
QII |
中更新世沉积物的冰复杂 |
砂壤土 |
|||
6.5 |
砂壤土 |
|||||
4/05 |
9.3 |
壤土 |
||||
4 |
科累马河低地,Chukochi斗篷 |
7/91 |
1 |
QIV |
的全新世沉积物唉 |
壤土与泥炭 |
科累马河低地,ole ltake |
1/95 |
1.25 - -1.3 |
壤土 |
|||
2/95 |
0.3 - -0.35 |
现代土壤 |
||||
科累马河低地, Kuropatochia河 |
2/96 |
10、6 - 10、7 |
QIII |
晚更新世沉积物的冰复杂 |
壤土 |
|
科累马河低地,科累马河河,Anuy河(Stanchikovsky和Duvanny纱线) |
露头1 |
土壤和洞穴埋在冰的晚更新世沉积物复杂 |
||||
露头2 |
||||||
分类群 |
种类和形式 |
||
CHROMALVEOLATA Adl et al . 2005 |
Alveolata Cavalier-Smith 1991 |
1852年纤毛亚门Doflein 1901 [Ciliata: Perty,纤毛虫类:Butschli 1887] |
1883年Oxytrichia sp。肾形虫属steinii Maupas肾形虫属inflata卡尔1931(1884年斯托克斯)肾形虫属aff. augustini Foissner 1987肾形虫属aff.aspera卡尔1926肾形虫属sp Platyophrya aff. vorax卡尔1926钟形虫sp。 |
隐藻纲采集1913年修订。Schoenichen 1925 |
1993年Goniomonadales Novarino和卢卡斯 |
1878年Goniomonas truncata(费森尤斯公司)斯坦 |
|
1989年Stramenopiles帕特森,修订。Adl等人。(2005) |
金藻纲采集1914 |
Spumella elongata(斯托克斯)贝尔彻和耗费1976 Spumella sp。 |
|
Incertae基准Alveolata |
Colponema edafi-cum Mylnikov et Tikhonenkov 2007 |
||
2002年EXCAVATA Cavalier-Smith修订。辛普森2003 (P ?) |
1985年Heterolobosea页面和布兰顿 |
Vahlkampfiidae Jollos 1917 |
Vahlkampfia sp。 |
Fornicata辛普森2003 |
Histionidae黄素和内拉德1993 |
Reclinomonas aff.美国黄素和内拉德1993 |
|
1981年Euglenozoa Cavalier-Smith修订。辛普森1997 |
眼虫目Butschli, 1884年修订。辛普森1997 |
Anisonema那Klebs 1893 |
|
Kinetoplastea Honigberg 1963 |
1914年博多curvifilis Griessmann博多designis Skuja 1948 b被Klebs 1893 b .小指Klebs 1893 |
||
1913年AMOEBOZOA Luhe修订。Cavalier-Smith 1998 |
Tubulinea斯米尔诺夫在Adl et al . 2005 |
Leptomyxida Pussard 1976年脑桥,修订。1987页 |
Leptomyxa sp。 |
索耶Acanthamoebidae脑桥1976年修订。1987页 |
Acanthamoeba sp。 |
||
Eumycetozoa Zopf, 1884年修订。橄榄1975 |
Incertae基准Eumycetozoa |
Hyperamoeba flagel-lata: Alexeieff 1923 |
|
(继续)
(继续)
分类群 |
种类和形式 |
||
Incertae基准AMOEBOZOA Spongomonadida:(Hibberd 1983)修订。卡尔波夫1990 |
Spongomonadidae卡尔波夫1990 |
1924年Phalansterium soli-tarium物质 |
|
Spongomonas uvella斯坦1878年 |
|||
1987年OPISTOCONTA Cavalier-Smith修订。Cavalier-Smith和超1995,修订。Adl et al . 2005 |
Choanomonada肯特1880 |
Monosigidae茹科夫和卡尔波夫1985 |
Codonosiga葡萄孢属肯特1880 |
Desmarella念珠肯特1880 |
|||
Monosiga ovata肯特1880 |
|||
Salpingoecidae肯特1880 |
Salpingoeca globulosa茹科夫1978年 |
||
RHIZARIA Cavalier-Smith 2002 |
1998年Cercozoa Cavalier-Smith修订。Adl et al . 2005 |
Cercomonadida (Poche 1913),修订。1983年Vickerman修订。Mylnikov 1986 |
Cercomonas angustus:(Skuja 1948) Mylnikov卡尔波夫2004 |
Cercomonas crassi-cauda雅1841 |
|||
Cercomonas granulif-era(奥朗德1942)Mylnikov卡尔波夫2004 |
|||
Cercomonas sp。 |
|||
Heteromita最小值(奥朗德1942)Mylnikov卡尔波夫2004 |
|||
1880年Heteromita aff. globosa肯特(斯坦) |
|||
Incertae基准Heteromitidae |
Allantion tachyploon Sandon1924 |
||
1990年Protaspis aff. gemmifera拉森和帕特森 |
|||
Protaspis单工V0rs 1992 |
|||
Incertae基准真核生物 |
Apusomonadida卡尔波夫和Mylnikov 1989 |
Apusomonadidae卡尔波夫和Mylnikov 1989 |
Apusomonas probosci-dea Alexeieff 1924 |
中阳目:库恩1926 |
Acanthocystidae老人1874 |
Choanocystis perpu-silla Siemensma 1991 |
|
-

- 图8.3光显微图的原生动物与冻土:a, b, c异养鞭毛虫;d, e, f裸体变形虫;g h,我纤毛虫。酒吧= 10 | im

大多数的物种都是bacteriotrophs,四种形式(Goniomonas truncata, Allantion tachyploon Colponema edaphicum, Choanocystis perpusilla)是捕食者。在安葬洞穴的一个样本,我们发现了一个centrohelid helio-zoan, Choanocystis perpusilla。对许多古代原生动物的物种,我们获得单一栽培和克隆文化,增长在20°C。
冻土样本收集3米以下表面(水井1/95,2/95,7/91,2/01,1/01,1/03,2/04,3/05,1/95)似乎更丰富的原生动物,因为他们是60%的样本中发现的。我们设法隔离的最大深度的原生动物是19 m(钻孔1/01)。生物发现的冻土上限不超过几百年,单身,最古老,发现可追溯至中更新世,200000 - 300000年(钻孔4/05;9.3米深)。
有一个倾向于可行的原生动物物种的数量和多样性埋土壤和洞穴更大比观察ice-complex沉积物。可能,这是解释cryoconservation更多有利条件和相对富裕的初始动物群埋土壤和洞穴。此外,收集
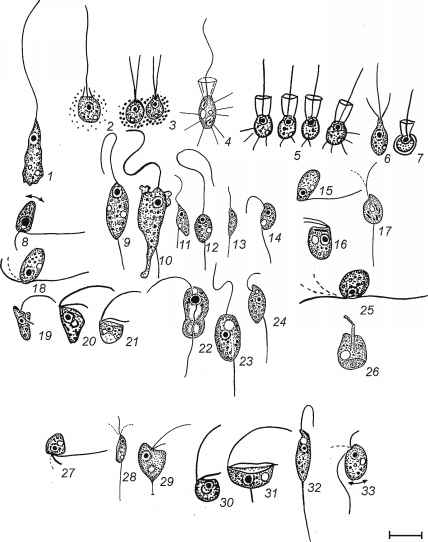
图8.4古异养鞭毛虫形态:1,Hyperamoeba鞭毛虫类;2,Phalansterium solitarium;3,Spongomonas uvella;4,Codonosiga葡萄孢属;5,Desmarella念珠;6,Monosiga ovata;7,Salpingoeca globulosa;8日,Cercomonas angustus;9日,Cercomonas crassicauda;10日,Cercomonas granulifera; 11, Cercomonas sp. 1; 12, Cercomonas sp. 2; 13, Cercomonas sp. 4; 14, Heteromita aff. globosa; 15, Allantion tachyploon; 16, Goniomonas trun-cata; 17, Protaspis aff. gemmifera; 18, Protaspis simplex; 19, 20, Spumella elongat; 21, Spumella sp. 1; 22, Colponema edaficum; 23, Anisonema ovale; 24, Bodo designis; 25, B. repens; 26, Apusomonas proboscidea; 27, Heteromita minima; 28, Cercomonas sp. 3; 29, Spumella sp. 2; 30, Spumella sp. 3; 31, Reclinomonas aff. americana; 32, Bodo curvifilis; 33, B. minimus. The bar is equal to 5 (1-27) or 10 (28-34) Jim
图8.4古异养鞭毛虫形态:1,Hyperamoeba鞭毛虫类;2,Phalansterium solitarium;3,Spongomonas uvella;4,Codonosiga葡萄孢属;5,Desmarella念珠;6,Monosiga ovata;7,Salpingoeca globulosa;8日,Cercomonas angustus;9日,Cercomonas crassicauda;10日,Cercomonas granulifera; 11, Cercomonas sp. 1; 12, Cercomonas sp. 2; 13, Cercomonas sp. 4; 14, Heteromita aff. globosa; 15, Allantion tachyploon; 16, Goniomonas trun-cata; 17, Protaspis aff. gemmifera; 18, Protaspis simplex; 19, 20, Spumella elongat; 21, Spumella sp. 1; 22, Colponema edaficum; 23, Anisonema ovale; 24, Bodo designis; 25, B. repens; 26, Apusomonas proboscidea; 27, Heteromita minima; 28, Cercomonas sp. 3; 29, Spumella sp. 2; 30, Spumella sp. 3; 31, Reclinomonas aff. americana; 32, Bodo curvifilis; 33, B. minimus. The bar is equal to 5 (1-27) or 10 (28-34) Jim of samples from outcrops allowed us to choose samples abundant in organics, more structured and, therefore, more suitable for protozoological examination.
没有透露相关可行的原生动物的发生之间的沉积物样品,这些沉积物的理化性质(水分、分级、pH值和温度)。
继续阅读:适应机制
这篇文章有用吗?