信息Xli
(1996)
卢塞恩
各种各样的草
作物
场
Bunce (1993)
大豆
反枝苋
作物
场
Ziska (2000)
*“青睐”表明是否升高[CO2]产生更多的作物和杂草生物量。“牧场”是指一种C3草地物种。
C4杂草(表2.2)。在这些比较,增加二氧化碳增加了作物杂草生物量比,符合已知的生化/手机/叶响应。然而,有趣的是指出,生物量和/或产量的高粱(C4作物)降低了高[CO2]在存在要么velvetleaf (Albutilon落叶或苍耳子(苍耳子strumarium), C3杂草。大多数比较相同的光合途径营养生长的作物和杂草导致作物和杂草生物量显著减少杂草和作物时同时出现(表2.2)。实际上只有两个研究量化作物与杂草丛生的种子产量竞争变化的函数上升[CO2] (Ziska、2000、2003 b)。在这些研究中,两种作物,C3(大豆)和一个C4(矮高粱),与藜生长(C3)和世界性(C4)和velvetleaf (C3)和世界性,分别在两个杂草的密度每米的行。从藜虽然大豆产量损失少,所有其他crop-weed交互导致增加产量损失升高[CO2]。有趣的是,在以后的研究中,任何杂草物种的存在否定的能力反应作物生长地或繁殖,增强[CO2]。这可能是重要的因为二氧化碳提高作物产量的研究很少考虑crop-weed竞争。然而,需要额外的实地研究证实并放大这里给出的结果。
2.7.2植物竞争:非托管系统
少即是已知的关于二氧化碳上升的影响植物之间的竞争非托管系统,在某种程度上,因为植物群落竞争涉及multispecies比较和最好的被认为是在一个生态系统上下文(参见2.8节)。此外,它可能很难单独升高的影响[CO2]等其他非生物资源的竞争水或营养。然而,有些情况下非托管系统中只有少数的物种竞争在给定的时间。例如,在早期,物种之间的竞争可以改变二氧化碳的函数(Bazzaz, 1996)。对森林系统、葡萄和增长缓慢的树表现出微分应对不断上升的二氧化碳,对葡萄树生物量与积极作用(例如格拉纳多斯& Korner, 2002)和后续影响葡萄树竞争(菲利普斯et al ., 2002)。C3和C4比较,高二氧化碳被证明有利于生物质生产的C3莎草(芦苇olneyi)生产的C4草(Spartinapatens)在湿地系统(Curtis et al ., 1989)。相比之下,主导C4草在主导青睐C3物种回应二氧化碳升高(Owensby et al ., 1993)在一个干燥,因为更高的高草草原系统干旱宽容的C4物种。
2.7.3二氧化碳改变植株相互作用如何?
毫无疑问,大气中的二氧化碳含量持续增加会改变植物竞争和组成(Bazzaz & McConnaughay, 1992)。考虑到经济和/或环境预测的竞争结果的重要性植物系统事实上,,我们知道的特定方面植物的生长和发展随着二氧化碳的增加与增加竞争成功?在细胞/叶水平我们可以认为二氧化碳的敏感性差C3和C4光合作用途径可以用来预测竞争的结果;然而,正如我们所见,这并不总是一个可靠的预测。此外,它并没有解决竞争的结果,光合作用途径是一样的。整个工厂层面我们可以认为,快速增长的物种更适应二氧化碳或营养压力提高二氧化碳反应;然而,这些也不是一个好的竞争结果的预测(例如要隘& Perez-Soba, 2001)。
目前,似乎没有任何“一刀切”的解释竞争成功应对二氧化碳。这不是太奇怪,因为竞争本身的复杂性。是一个伟大的需要详细的植株相互作用的研究,有助于解决特定的时间和空间开发地上(如光)或地下的(如氮、水)资源来应对二氧化碳的增加。
2.8植物群落和生态系统对二氧化碳的反应
2.8.1发布工厂管理系统
因为的重要性食品安全,更早期的关于二氧化碳上升的影响的重点是农作物(例如歪斜着&艾伦,1985)。然而,许多这些个体植物的研究,和实地评估作物系统只是明显从1990年代(例如金博et al ., 1995)。一般来说,二氧化碳浓度200 - 400 ppm(通常高于当前的环境水平)已发现刺激增长和产量C3(大米、小麦),但不是C4谷物(玉米、高粱);并刺激豆科结节作物(大豆)和(土豆)以及许多绿叶蔬菜(审查看到Reddy &霍奇斯,2000)。纤维作物,如棉花,也可以显示出强劲增长和棉子回应二氧化碳升高(金伯尔& Mauney, 1993)。另外,牧场并不总是表现出强烈回应二氧化碳(例如Korner, 1997)。
尽管年度作物响应管理系统已经深入研究,少即是已知的关于管理的物种。世界上森林种植园现在总约130尼古拉斯年利率约10.5尼古拉斯(詹森的建立等等。,2000)。在美国,商业种植火炬松(松果体taeda)仍然是一个木材产品的主要来源(Jokela et al ., 2004)。而成熟的泥泞的反应已经在应对二氧化碳在非托管系统检查(例如DeLucia et al ., 1999),培养和受精的反应是未知的。同样,CO2-induced石头或热带水果的生产力的变化在很大程度上未经检验的(詹森et al ., 2000)。
2.8.2用水管理系统
气孔和叶地区回应二氧化碳升高有最好的特点是年度作物(Bunce, 2004 b)。但单一叶反应转化为减少用水在社区一级管理系统?发育,增加二氧化碳会导致增加叶面积和工厂规模以及R / S比值的变化,植物解剖学、和导电组织拍摄的增长(批&亚历山大,1993)。因此,目前尚不清楚是否有水分或增加的储蓄叶作物群体内观察到的水平。
虽然作物中最大的降低气孔导度在升高[CO2],相对日常减少与其他非生物输入的不一致,可能相差很大(如光、温度)和广泛的气孔可变性(Bunce, 2004)。总的来说,使用模拟和直接测量系统,任何大型减少作物气孔导度只会翻译成小减少社区蒸散,在某种程度上,因为直接影响二氧化碳的增加冠层温度和减少湿度(Bunce, 2004)。这些后来的变化也有重要的对作物产量的影响(如松井et al ., 1997)。然而,在系统层面,甚至降低几个百分点的蒸散可能是重要的作物产量和作物生产的经济。
2.8.3 Unmanagedplant系统
方法论的变化,特别是面对技术的出现在1990年代,激发兴趣解决二氧化碳上升的潜在影响在社区层面的反应(Hendrey & Kimball, 1994)。然而,非托管系统的响应是复杂的,因为不同农业管理,非生物输入例如水或营养物质可以是非常变量。尽管非托管系统可以显示增加生产力和植物物种组成的变化在回应二氧化碳升高(如史密斯等等。,2000年),有一个广泛的具体预测二氧化碳升高将如何改变社区层面的过程。
二氧化碳反应的早期评估在北极苔原的系统中,例如,表现出生产力的变化不大(Grulke et al ., 1990)。草原社区显示混合增长反应[CO2],与社区更大程度的物种丰富度显示更大的反应(例如帝国et al ., 2001),可能由于高二氧化碳反应物种少没有出现在不同的社区(例如Grunzweig & Korner, 2000)。沙漠生态系统,高架(二氧化碳)影响的程度是与降雨(即增加水分和养分)与后续增加社会生产力(史密斯et al ., 2000)。相反,植物中湿地系统(如s olneyi马什C3物种)继续显示特有的正增长回应二氧化碳升高17年后的暴露(麝香猫et al ., 2005)。
在非托管系统,二氧化碳上升的影响森林生产力是特别感兴趣的,鉴于森林在陆地碳汇的作用。一般来说,回顾与小树的长期实验也表明显著增加(~ 30%)增长一倍[CO2]从目前水平(Medlyn et al ., 2001)。然而,目前尚不清楚,得到的差异大气候raybet雷竞技最新开启和关闭之间的树冠上,类似的反应是否会被观察到暂时的增长和净初级生产力(NPP)的更成熟。到目前为止,大部分的实验证据表明,可能有一个永久性的二氧化碳的影响即使出现光合适应,但是额外的信息,特别是在地下的碳分配(Zak et al ., 2003)是必要的。
2.8.4水使用的非托管植物系统
草原和沙漠,有明显的间接相互作用(气孔)[CO2]对用水量的影响和刺激植物生长在社区一级。摩根et al。(2004)最近回顾一些田间试验包括堪萨斯的高草草原,科罗拉多shortgrass草原,和瑞士钙质草原、所有系统显示更大的干年[CO2]增强。相比之下,德州C3 / C4草原和新西兰牧场每年影响土壤水分的变化,而在莫哈韦沙漠只是刺激植物生长在湿年升高[CO2]。虽然之间的交互(CO2)和水的可用性是明显在这些生态系统,到目前为止,没有系统性的二氧化碳分离浓缩反应与间接推动反应已经完成实验(Morgan et al ., 2004)。同样,没有长期的评估分离CO2施肥从用水可用于森林群落的影响。长期水文平衡评估为公爵的脸设施表明火炬松没有直接影响的高二氧化碳在水储蓄可辨别的(3.5年);相反,森林逐渐发生更多的水,可能由于减少土壤蒸发由于额外的垃圾堆积在高[CO2](谢弗等等。,2002)。
2.8.5其他营养级
任何考虑生态系统响应增加[CO2]应该不仅包括植物生产力,但也潜在影响高营养水平。例如,它可能将影响食草动物生物学的高二氧化碳对寄主植物代谢的生理效应。具体CO2-induced变化在叶级将包括C / N比值增加,改变防御化合物的浓度,增加了淀粉和纤维含量,增加水分含量(例如林肯& Couvet, 1989)。不太清楚,但是,是否响应观察到叶或植物水平与植物群落的反应是一致的。例如,有补充叶产量的变化,可能,可能,克服insect-related损害(休斯& Bazzaz, 1997)。矮橡树和湿地生态系统,以树叶为食的动物的侵扰观察到二氧化碳升高(汤普森&德雷克,1994;阶梯et al ., 2002)。近期数据对舞毒蛾在成熟森林表明种特异的叶化学成分的变化引起的高[CO2]可能导致对比食草动物反应(Hatten-schwiler & Schafellner, 2004)。优惠食草动物饲养在一个物种可能,反过来,改变植物的竞争。然而,总的来说,大多数数据只进行单一insect-host植物相互作用来应对二氧化碳的增加,和一个更完整的评估缺乏植物群落内的昆虫取食。
也有许多公认CO2-induced变化可能会改变植物对疾病的易感性。例如,提高WUE和叶片水分含量可以促进由叶面真菌孢子形成(例如汤普森&德雷克,1994),同时增加叶碳水化合物能促进病原体感染后的生长和繁殖(Hibberd et al ., 1996)。另外,减少叶片含氮量可以减少病原体负载和疾病严重程度(汤普森et al ., 1993)和CO2-induced衰老变化可以增加或减少暴露时间对病原体(例如表示&领域,1997)。在社区层面,减少蒸发蒸腾作用也可以减少与顺向树冠湿度对大多数真菌的生长和孢子形成的影响(Chakraborty et al ., 2000;Chakraborty &数据,2003),而提高生产率高[CO2]可以增加植物残体,与更大的潜在致病过冬(Manning &蒂德,1995)。此外,根产量的增加和/或根分泌的变化会增加宿主组织用于病原感染的比例(Manning &蒂德,1995)。然而,总的来说,极其有限的关注这一领域的研究排除了任何使广义预测与信心的能力。剩下的,而一般预测的疾病可能会增加,减少,或显示没有变化”(科克利,1995)。
2.9全球和进化鳞片
2.9.1上升二氧化碳作为选择的因素
使用图2.2作为指导检查二氧化碳上升改变了植物生物功能如何在时间和空间,我们没有考虑限制沿着轴。例如,似乎不太可能,植物或社区应对二氧化碳将随着时间的推移保持稳定;然而,很少有人注意的后果增加二氧化碳在进化的时间尺度。与光、营养物质和水,有相当大的遗传变异,以应对二氧化碳(例如柯蒂斯et al ., 1994;Bazzaz et al ., 1995),表明植物繁殖和进化改变成功。在较短的时间尺度,评价这些选择性变化可能务实的后果,如高产农艺栽培品种的选择在工厂管理系统(如安斯沃思et al ., 2002)或入侵植物物种的成功在一个社区(史密斯et al ., 2000;Hattenschwiler & Korner, 2003)。长期评估物种的成功,目前生物多样性的变化,或社区选择高营养水平以应对二氧化碳本身是不可用。
2.9.2全球影响
如果进化是一个长期的时间改变,那么全球估计关于二氧化碳上升的影响生态系统功能反映了一个非常大的空间。如此规模的改变不能解决实验,但只有通过全球建模,以及大气环流模型以及区域气候评估是可用的,包括atmosphere-biosphere交流(如布尔et al ., 2000)。raybet雷竞技最新这样的模型集成和综合现有的信息关于二氧化碳的影响和全球项目信息的结果。因此,建模工作是有用的作为潜在全球后果的预测,并强调地方附加查询是必要的。
然而,考虑到规模大的调整,不确定性在生态系统水平在全球评估大幅放大。举个例子,有一个很大的兴趣量化植物群落在隔绝额外的碳的作用作为一个潜在的手段减少大气中的二氧化碳增加的速度(如格尼et al ., 2002)。在社区/生态系统水平上,有一些问题,森林系统的能力作为长期碳汇(例如施莱辛格&里胥特博士,2001),强调需要更好地了解碳/氮循环在森林土壤。事实上,土壤氮池的作用显得至关重要的理解全球碳封存和仍然大量讨论的主题(例如Norby & Cotrufo, 1998;Zak et al ., 2003;Hungate et al ., 2004年,参见章节8和9)。然而,全球气候和建模估计CO2-induced NPP的增加碳封存(以及随后的变化)(例如Nemani raybet雷竞技最新et al ., 2003)可能不会考虑这样的微妙之处。不幸的是,政策制定者常常认为气候变化模型最后,权威的评价,而不是在进步。raybet雷竞技最新
2.10的不确定性和局限性
二氧化碳是植物生物学四个必要的非生物输入和最近和预测它的浓度的变化已经影响,并将继续影响,植物功能。虽然二氧化碳的主要生理作用是直接关系到碳吸收/损失和用水,很明显,这些变化改变植物功能在每一个组织层面(图2.3)。同样清楚的是,一个实验性的或概念的理解在一个组织层面未必作为一个可靠的预测各级功能行为的指南。全面掌握叶级流程,例如,只提供有限的了解生态系统响应。然而太多的关于二氧化碳对植物生物学的影响仍然是描述性的,而不是机械的;专注于单一植物响应,non-integrative。总的来说,在评估植物对二氧化碳的反应时,有明显的人员必须扩大他们的发现。
组织需要更多的实验研究水平?虽然有很多评价,检验个体植物的光合作用和生长反应(二氧化碳)的两倍,有相对较少的报道关于二氧化碳上升的影响在空间或时间极端。例如,我们知之甚少特定CO2-induced基因表达的变化,这些变化将如何影响或一个进化上的意义;同样,我们几乎一无所知的二氧化碳长期生态系统功能的影响和碳之间的交互,水和养分循环(如锅et al ., 1998)。
但最终,生态系统集成商的基因、结构,能辨出别交互,较高的营养级,进化响应在时间和空间。此外,这些影响,造成全球增加大气中的[CO2],无论经济或环境,是最有可能被证明在生态系统功能。对于管理系统,增加二氧化碳的作用不受欢迎的植物的成功,尤其是入侵或有害物种,值得特别关注;在非托管系统中,微分物种的反应,以及由此产生的对物种选择、多样性、和更高的营养水平,仍然是一个重要的领域
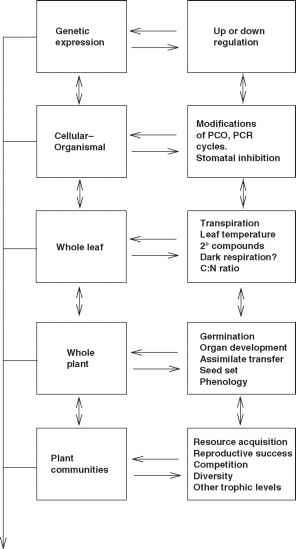
图2.3增加大气中的二氧化碳的潜在影响不同组织层次如图2.2所示。更多细节给出了文本。
生态系统功能
图2.3增加大气中的二氧化碳的潜在影响不同组织层次如图2.2所示。更多细节给出了文本。
研究。有明显的实验挑战研究生态系统(如非生物变迁和同比初级生产力的变化,量化地下过程尤其是养分循环和碳储存,潜在的食草性的变化,病原体负载,等等);然而,这些假设,考虑多因素的反应,特别是在生态系统层面,是必要的,如果我们要明确认识和适应CO2-induced植物系统的变化。
还有另外一个基本的挑战:全球二氧化碳的增加需要认识到,仅仅是前所未有的人为改变的一个方面。目前有60亿人口,人类是显著改变氮沉积率(例如Wedin & Tilman, 1996),对流层臭氧的程度(例如Krupa &曼宁,1988),和土地使用模式(皮尔克et al ., 2002)。任何关注生态系统动力学的实验方法,因此,应该考虑不仅二氧化碳还其他快速变化的非生物变量,只要有可能。希望多因素方法集成生态系统功能还可以用来增加现有的全球变化模型的预测能力。
引用
翘起的,b &艾伦L.H.小(1985)作物反应高浓度二氧化碳浓度。:增加二氧化碳的直接影响植被(eds开国元勋之一B.R.安贝德卡对应变& J.D.治愈),53 - 98页。美国能源部、美国能源部/ er - 0238,华盛顿特区亚当,无危险墙,G.W.金博,文学士学位品特,P.J.小,LaMorte R.L.、Hunsaker D.J.、Adamsen F.J.汤普森,T。马提亚,公元莱维特,S.W.& Webber, A.N. (2000) Acclimation response of spring wheat in a free-air CO2 enrichment (FACE) atmosphere with variable soil nitrogen regimes. 1. Leaf position and phenology determine acclimation response. Photosynth. Res., 66, 65-77.
安斯沃思,电子艺界戴维,公共广播、Bernacchi C.J.Dermody,超频希顿,电子艺界,摩尔。D.J.摩根,P.B.,Naidu S.L.Yoo H.-S。、朱、X.G.柯蒂斯,注:&长……(2002)[CO2]升高影响的荟萃分析大豆(大豆)生理学、生长和产量。全球变化生物学观点》,695 - 709。
安斯沃思,电子艺界戴维,公共广播、Hymus G.J.奥斯本,C.P.罗杰斯。布卢姆,H。Nosberger, j . &长……(2003)是由二氧化碳浓度升高的刺激叶光合作用维持长期的吗?测试与多年生黑麦草种植10年两个氮肥水平下自由空气二氧化碳浓缩(脸)。植物细胞包围。26, 705714.
阿尔贝托,点Ziska, L.H.、Cervancia C.R. & Manalo公共广播(1996)增加二氧化碳和温度的影响有竞争力的相互作用之间的C3作物,水稻(栽培稻)和C4杂草(Echinochloa glabrescens)。欧斯特。j .植物杂志。,23岁,795 - 802。Amthor, j.s(1994)扩展CO2-photosynthesis树冠从叶的关系。Photosynth。Res, 39岁,321 - 350。
Amthor, j.s科赫,G.W.,Williams jr &Layzell D.B. cooper(2001)叶O2吸收在黑暗中独立于重合的二氧化碳分压。j . Exp。机器人。,52岁,2235 - 2238。Arp, j(1991)源库关系的影响光合适应二氧化碳升高。植物
细胞包围。,869 - 875。Assman,克里(1999)保卫细胞的细胞基础传感上升的二氧化碳。植物细胞包围。22, 629-638.
Bae, r·h·&西歇尔(2004)可溶性蛋白表达的变化,叶子代谢物水平Ara-bidopsis芥生长在大气二氧化碳浓度升高。田间作物Res, 90, 61 - 73。
Bazzaz, 11(1996)植物在变化的环境中:链接生理、人口和社会生态。英国剑桥大学出版社、剑桥。
Bazzaz,碰头Jasienski, M。、托马斯、南卡罗来纳州和韦恩,p(1995)微观进化反应实验的植物种群CO2-enriched环境:平行结果从两个模型系统。Proc。国家的。学会科学。美国,92年,8161 - 8165。
Bazzaz,粮农组织& McConnaughay K.D.M.(1992)能辨出别在高二氧化碳环境中相互作用。欧斯特。j .机器人,547 - 563。
比林斯,S.A.Zitzer,旧金山那里Weatherly, H。Schaeffer,克里查莱特,T。Arnone,正当三世&埃文斯,r(2003)升高的二氧化碳对落叶层质量的影响在莫哈韦沙漠生态系统。全球变化生物学观点》,729 - 735。
布尔G.J.弗拉托,通用,读者,m &冉斯登,d .(2000)瞬态模拟与历史和预测气候变化的温室气体raybet雷竞技最新气溶胶强迫:实验设计和与20世纪的仪器记录。爬。直流发电机,16岁,405 - 425。
Bowes, g(1996)光合反应大气二氧化碳浓度的变化。:光合作用与环境(ed。天然橡胶贝克),387 - 407页。血案》出版,荷兰多德雷赫特。
Bunce, J.A.(1993)增长、生存竞争,树冠二氧化碳和水汽交换第一年苜蓿在二氧化碳浓度升高。Photosynthetica, 29岁,557 - 565。
Bunce, J.A.(1994)反应呼吸的大气二氧化碳浓度的上升。杂志。工厂。,90,427 - 430。
Bunce j (1995 a)在黑暗中二氧化碳浓度升高对大豆幼苗的生长。安。机器人。,75,365 - 368。
Bunce j (1995 b)长期发展苜蓿和果园草块二氧化碳升高。j . Biogeochem。,22岁,341 - 348。
Bunce, J.A.(2000)适应环境温度在8个酷和光合作用温暖的气候raybet雷竞技最新草本C3物种:生化参数的温度依赖光合作用模型。Photosynth。Res, 63, 59 - 67。
Bunce, J.A.(2001)直接和acclimatory反应气孔导度升高的二氧化碳在四个草本作物物种。全球变化生物学观点》,323 - 331。
Bunce j (2004 a)二氧化碳影响气孔对环境的反应和水作物田间条件下使用。环境科学,140年1 - 10。
Bunce j (2004 b)比较二氧化碳浓度和温度对呼吸的影响,易位和硝酸还原黑大豆叶子。安。机器人。,93,665669。
Bunce j &自信司令部(2001)水的压力和日常field-grown大豆的光合适应环境的变化,二氧化碳浓度升高。Photosynthetica, 39岁,95 - 101。
Bunce j &自信司令部(2003)日常辐照度和反馈抑制光合作用在甘蓝oleracea二氧化碳浓度升高。Photosynthetica, 41岁,481 - 488。
卡特,湄&彼得森K.M. (1983) CO2-enriched气氛对经济增长的影响和竞争互动的C3和C4草。环境科学,58岁,188 - 193。
Chakraborty s &数据,美国植物病原真菌(2003)将如何适应寄主植物抵抗气候变化下二氧化碳升高?raybet雷竞技最新新植醇。,159,733 - 745。
Chakraborty, S。、Pangga I.B.勒普顿,J。哈特,L。房间,下午&耶茨d(2000)生产和传播Stylosanthes刺盘孢属、炭疽孢子的作用下二氧化碳升高。环绕。Pollut。,108,381 - 387。
程、工程学系。摩尔,最初& Seemann jr(1998)的影响短期和长期的二氧化碳Ribulose-1表达的升高,5-bisphosphate羧化酶/加氧酶基因在拟南芥的叶子和碳水化合物的积累(l)Heynh。植物杂志。,116,715 - 723。
基督,R.A. & Korner c(1995)拍摄和根的反应气体交换,叶片扩张和生物质生产的脉冲二氧化碳升高水培小麦。j . Exp。机器人。,46岁,16611667。
楚,贝科尔曼,J.S.二&穆尼H.A.(1992)之间的生物质分区控制根和拍摄:大气二氧化碳浓缩和碳和氮的获取和分配野生萝卜。环境科学,89,580 - 587。
科克利,克里(1995)生物圈的变化:在植物病理学,会有什么关系吗?可以。j .植物病理学研究。,17岁,147 - 165。
表兄弟,学士亚当,天然橡胶墙,G.W.金博,文学士学位品特,P.J.、小Ottman, M.J.莱维特,S.W.& Weber, A.N. (2003) Development of C4 photosynthesis in sorghum leaves grown under free-air CO2 enrichment (FACE). J. Exp. Bot., 54, 1969-1975.
柯蒂斯,注:德雷克,人Leadley,毛重Arp, j & Whigham测向(1989)生长和衰老的植物群落暴露在二氧化碳浓度升高河口湿地。环境科学,78,20-26。
柯蒂斯,注:,雪,&米勒A.S. (1994) Genotype-specific二氧化碳升高对繁殖力的影响在野生萝卜获得raphanistrum)。环境科学,97,100 - 105。
柯蒂斯,注:& Wang x(1998)升高的二氧化碳影响的荟萃分析木本植物质量,形式和生理学。环境科学,113,299 - 313。
戴维,公共广播亨特,S。、Hymus G.J.DeLucia,再见德雷克,人、Karnosky测向&长……(2004)呼吸耗氧量不减少二氧化碳的瞬时高程,但增加在高二氧化碳领域长期发展。植物杂志。,134,520 - 527。
DeLucia,再见,汉密尔顿j。,ShawnaL.N。托马斯·R.B.,安德鲁斯正当,Finzi A。Lavine M。Matamala, R。莫汉,J.E.Hendrey, G。R. & Schlesinger, W.H. (1999) Net primary production of a forest ecosystem with experimental CO2 enrichment. Science, 284, 1177-1179.
德雷克,人Azcon-Bieto, J。浆果,正当Bunce,正当迪杰斯特拉,P。法勒,J。科赫,G.W.琥珀,H。,j . & Wullschleger Siedow s(1999)二氧化碳升高抑制植物线粒体呼吸在绿色植物吗?植物细胞包围。22, 649-657.
德雷克,人Gonzalez-Meler, m &长……(1997)更高效的植物:二氧化碳上升的后果吗?安。启植物杂志。植物摩尔。生物。现年48岁的609 - 639。
德雷克,人& Leadley毛重(1991)树冠作物的光合作用和本地植物群落长期暴露在高二氧化碳。植物细胞包围。14, 853-860.
庄士贤,点&诺贝尔,注:(2000)凸轮物种的反应增加大气中的二氧化碳浓度。植物细胞包围。23, 767-781.
曼森Ehleringer jr &上面(1993)光合途径变异的进化和生态方面。安。启生态。系统。,24岁,411 - 439。
Esashi Y。安倍,y & Ashino h(1989)苍耳属植物种子的萌发和生长的轴向和子叶的组织对乙烯反应,二氧化碳,氧气和/或在水下的压力。植物细胞包围。12, 183-190.
Esashi Y。,哈泽,s &小岛,k(1987)光行动苍耳属植物种子的萌发:诉当地乙烯,二氧化碳,氧气在发芽的关系。j . Exp。机器人。,38岁,702 - 710。
Ferrario-Mery, S。、Thidaud M.-C。Betsche, T。艾尔丁M.-H。&门厅,h(1997)调制的碳和氮代谢和硝酸还原酶,untransformed烟草plumbagini-folia在二氧化碳浓缩锅和水培的植物生长。足底、202、510521。
Garbutt, k & Bazzaz碰头(1984)升高的二氧化碳对植物的影响。三世。花、水果和种子生产和堕胎。新植醇。,98,433 - 446。
Garbutt, K。& Bazzaz威廉姆斯、11(1990)的微分响应分析高浓度二氧化碳五一年生植物在增长。生态学,71,1185 - 1194。
盖革,M。Haake, V。Ludewig F。,美国&施迪Sonnewald m(1999)硝酸和硝酸铵供应产生重大影响光合作用的反应,碳代谢、氮代谢和生长在烟草二氧化碳升高。植物细胞包围。22, 1177-1199.
盖革,M。Walch-Piu, L。Harnecker, J。Schulze工作流管理。Ludewig F。Sonnewald U。、Scheible W.-R。&施迪m (1998)。增强二氧化碳导致改性硝酸还原酶活动的昼夜节律老工厂,和一个大的模拟硝酸还原酶活性和更高水平的氨基酸在高等植物。植物细胞包围。21, 253-268.
Ghannoum, O。,冯·Caemmerer年代。Ziska, L.H.& Conroy, J.P. (2000) The growth response ofC4植物大气中的二氧化碳分压上升:重新评估。植物细胞包围。23, 931-942.
Gonzalez-Meler,硕士Ribas-Carbo, M。Siedow J.N. &德雷克,人(1996)直接抑制植物线粒体呼吸二氧化碳升高。植物杂志。,112,1349 - 1355。
Gonzalez-Meler,硕士Taneva L & Trueman R.J.(2004)植物的呼吸作用和大气中二氧化碳浓度升高:细胞反应和全球意义。安。机器人。,94,647 - 656。
格拉纳多斯j . & Korner c(2002)在树荫深处,二氧化碳会增加热带攀缘植物的活力升高。全球变化生物学观点》,1109 - 11雷竞技csgo17。
Grulke, N.E.瑞切尔斯,G.H.Oechel,后来,美国& Jaeger Hjelm c(1990)在草丛苔原碳平衡环境和大气中的二氧化碳升高。环境科学,83,485 - 494。
Grunzweig, j . & Korner c(2000)生长和生殖回应二氧化碳升高北部以色列内盖夫的野生谷物。全球变化生物学观点》6,631 - 638。
格尼,投资者、法律、智慧化丹宁,响亮的雷纳,P.J.贝克,D。Bousquet, P。Bruhwiler, L。陈,中州。Ciais, P。粉丝,S。Fung, I.Y.Gloor, M。这个,M。Higuchi, K。,约翰,J。真希,T。、Maksyutov年代。Masarie, K。Peylin, P。普莱瑟,M。Pak,公元前安德森,J。Sarmiento, J。田口方法,S。高桥,t &袁C.-W。 (2002) Towards robust regional estimates of二氧化碳的源和汇使用大气传输模型。自然,415,626 - 630。
哈代,R.W.F. & Havelka U.D.K.(1975)共生N2固定二氧化碳增多:全方位的增强的场地大豆。植物杂志。现年48岁的35。
Hattenschwiler, s & Korner c(2003)二氧化碳升高促进入籍的沙卡李属laurocerasus的瑞士温带森林?功能。生态。,17岁,778 - 785。
Hattenschwiler, s & Schafellner c(2004)舞毒蛾喂养的树冠二氧化碳丰富成熟的森林。全球变化生物学观点》,1899 - 1908。
Hendrey,广义相对论& Kimball,文学士学位(1994)程序。阿格利司。对。Meteorol。,70年,3 - 14。
Hibberd, J.M.惠特布莱德,r &法勒,参考书籍(1996)效应的二氧化碳浓度的升高对由白粉菌属感染大麦小麦。杂志。摩尔。植物病理学研究。现年48岁的37-49。
Holtum J.A.M. &冬天,k(2003)幼苗的光合作用吸收二氧化碳热带的树物种暴露于振动的二氧化碳浓度升高。足底,218,152 - 158。
霍顿,j.t丁,Y。、Griggs D.J.Noguer, M。范德林登,P.J. & Xiaosu d (eds)(2001) 2001年气候变化:科学依据。raybet雷竞技最新英国剑桥大学出版社、剑桥。
休斯,l . & Bazzaz碰头(1997)升高的二氧化碳对相互作用的影响西方花蓟马,Frankliniella occidentalis(缨翅目:蓟马科)和常见的马利筋,Ascelias syriaca。环境科学,109,286 - 292。
Hungate,文学士学位阶梯,下迪杰斯特拉,P。约翰逊,D.W.凯特,工程师、Hymus G.J.亨克尔,C.R. &德雷克伞齿轮(2004)二氧化碳抒发长期下降固氮。科学,304,1291。
Idso, C.D.、Idso S.B. &球团司令部小(1998)城市二氧化碳圆顶的凤凰城,亚利桑那州。理论物理。地理,19岁,95 - 99。
雅布隆斯基,L.M.王,x &柯蒂斯,注:(2002)植物繁殖高二氧化碳条件下:一个荟萃分析报告79作物和野生物种。新植醇。,156年,9-26。
Jach卷& Ceulemans r(1999)大气中的二氧化碳升高对物候学的影响,增长和皇冠欧洲赤松的结构(抗旱性)幼苗经过两年的接触。树杂志。,19岁,289 - 300。
杨克,s & Krewitt m(2002)大气中的二氧化碳浓度会直接影响烟草的叶子呼吸测量,但不是呼吸本身。植物细胞包围。25, 641-651.
詹森,i。摩梭,m & Ceulemans r(2000)作物生态系统对气候变化的反应:树作物。:气候变raybet雷竞技最新化和全球作物生产力(eds投资者Reddy & H.F. Hodges), 245 - 263页。CABI出版社,纽约。
贾维斯P.G. & McNaughton kg。(1986)气孔蒸腾的控制:扩大fromleaf地区。放置生态。15日,1-49 Res。
Jokela, E.J.多尔蒂,下午和马丁,助教(2004)生产动力学ofintensively管理火炬松站在美国南部:七个长期实验的合成。对。生态。管理。,192,117 - 130。
h·g·琼斯(1998)气孔控制光合作用和蒸腾作用。j . Exp。机器人。现年49岁,387 - 398。
卡普兰,A。盖尔,J。& Poljakoff-Mayber, A. (1977) Effect of O2 and CO2 concentrations on gross dark二氧化碳的固定和暗呼吸在Bryophyllum daigrmontianum。欧斯特。j .植物杂志。4,745 - 752。
Karnosky,测向Ceulemans, R。Scarascia-Mugnozza, ge & Innes J.L. (eds)(2001)二氧化碳和其他温室气体的影响森林生态系统。CABI出版社,瓦林福德,英国。
Kerslake, J.E.,中华民国与哈特利Woodin S.E.(1998)二氧化碳和氮的影响浓缩plant-insect交互:质量Calluna寻常的作为东道主为Operophtera brumata。140年NewPhytol 43-49。
金博,学士(1993年)增加大气中的二氧化碳对植被的影响。Vegetatio 104/105 65 - 83。
金博,文学士学位& Mauney jr(1993)反应的棉花不同二氧化碳,灌溉和氮:产量和增长。阿格龙。J。,85,706 - 712。
金博,文学士学位品特,P.J.,加西亚,R.L. Jr、LaMorte R.L.墙,G.W.、Hunsaker D.J.Wechsung, G。,f . & Kartschall Wechsung t(1995)下的小麦生产力和用水fee-air二氧化碳浓缩。全球变化生物学观点》,429 - 442。
科赫,G.W. &穆尼H.A.(1996)二氧化碳和陆地生态系统。学术出版社,圣地亚哥,CA。
科赫,动向(1996)Carbohydrate-modulated在植物基因表达。安。启植物杂志。植物摩尔。生物。,509 - 540。
Korner, c(1997)的反应高山草原四季的二氧化碳增多:合成。Acta Oecol。,18岁,165 - 175。
两部,。霍夫曼,B。谢弗,c &施迪m(1993)调节红细胞表面的表达由碳水化合物和其他光合基因:光合作用的“沉”监管机制?植物J。,817 - 828。
Krupa S.V. &曼宁j(1988)大气臭氧:形成和对植被的影响。环绕。Pollut。,101 - 137。
Lavola, a & Julkunen-Titto r(1994)升高的二氧化碳和受精的影响在初级和次级代谢物桦木、桦木属翻车机(罗斯)。环境科学,99,315 - 322。
刘易斯,J.D.王,X.Z.格里芬,K.L. &组织D.T.(2002)年龄和个体发生对光合作用的影响反应的决定性一年生植物二氧化碳浓度升高。植物细胞包围。25, 359-368.
林,W。Ziska, L.H.Namuco运行系统&白,k(1997)交互的高温和高二氧化碳单叶片的光合适应水稻原位。杂志。工厂。,99,178184。
林肯,D.E. & Couvet d(1989)的影响碳供应分配物质和卡特彼勒食用薄荷。环境科学,78,112 - 120。
Lindroth R.L. & Kinney,株式会社(1998)的后果丰富大气中的二氧化碳和落叶叶的化学和舞毒蛾的性能。j .化学。生态。,24岁,1677 - 1689。
Lindroth, R.L.Kinney,株式会社&广场建筑中一段(1993)的反应落叶乔木大气中的二氧化碳升高:生产力、植物化学和昆虫的性能。生态学,74,763 - 772。
劳埃德,j . &法夸尔,国民生产总值(1996)的二氧化碳依赖光合作用,植物生长对大气CO2浓度升高的反应及其与土壤养分状况。我一般原则和森林生态系统。功能。生态。10 4-32。
长,……(1991)修改光合生产力的反应温度升高,大气中的二氧化碳浓度:它的重要性被低估了吗?植物细胞包围。14, 729-739.
Ludewig f & Sonnewald,(2000)高CO2-mediated下调光合基因转录是由加速叶片衰老引起的而不是糖积累。2月。,479年,19到24。
罗,y &穆尼H.A. (eds)(1999)二氧化碳和环境压力。学术出版社,圣地亚哥,CA。
牧野,A。Nakano, H。美,T。岛田t &山本,N(2000)光合作用,植物生长和N分配在转基因水稻与降低二磷酸核酮糖羧化酶在二氧化碳浓缩。j . Exp。机器人。,50383 - 389。
表示:&,C.B.(1997)占据不同的燕麦植物高浓度二氧化碳的反应。植物细胞包围。20, 178-185.
曼宁,j &蒂前任所长A.V.(1995)气候变化:增加大气二氧化碳(CO2)raybet雷竞技最新的潜在影响,臭氧(O3)紫外线b(uv - b)辐射对植物病害。环绕。Pollut, 88, 219 - 234。
Masle, j .(2000)二氧化碳浓度升高的影响细胞分裂率,增长模式,和年轻的小麦叶片解剖植物叶的位置是通过控制调节因素相关,春化和基因型。植物杂志。,122,1399 - 1416。
松井,T。Namuco,运行系统Ziska, L.H.& Horie, T. (1997) Effects of high temperature and CO2 concentration on spikelet sterility in Indica rice. Field Crops Res., 51, 213-221.
Medlyn, B.E.雷伊,。巴顿,C.V.M. & Forstreuter m(2001)地上林木的生长反应,大气中的二氧化碳浓度升高。:二氧化碳和其他温室气体的影响森林生态系统(eds测向Karnosky, r . Ceulemans ge Scarascia-Mugnozza & J.L. Innes), 127 - 146页。CABI出版社,纽约。
宫崎骏,S。弗雷德里克森,M。霍利斯,kcProyko, V。Shepley D。加尔布雷斯,D.W.、长……& Bohnert H.J.(2004)转录表达谱的拟南芥受控条件下的生长和露天CO2和O3的浓度升高。90年田间作物Res, 47-59。
莫汉,J.E.陶伟洪,克拉克,j.s &施莱辛格(2004)遗传变异在发芽,成长和生存的红枫在回应subambient通过大气中的二氧化碳升高。全球变化生物学观点》,233 - 247。
摩尔,最初程学系。西姆斯,d & Seemann jr(1999)的生化和分子基础光合适应大气中的二氧化碳升高。植物细胞包围。22, 567-582.
摩根,正当亨特,H.W.,c.a & LeCain Monz湄(1994)增长的后果在两个二氧化碳浓度和两个温度下的叶片气体交换Pascopyrum smithii (C3)和Bouteloua股薄肌(C4)。植物细胞包围。17, 1023-1033.
摩根,正当帕塔基,D.E.Korner C。克拉克,H。德尔格罗索,中华民国、Grunzweig J.M.克纳普,A.K.那么,Mosier境牛顿,P.C.D.Niklaus,公共广播、Nippert J.B.诺瓦克,R.S.帕顿,j•、波莉·&肖,核磁共振(2004)水关系在草原和沙漠生态系统暴露在大气中的二氧化碳升高。环境科学,140,11-25。
莫里森,J.I.L.(1998)气孔响应增加二氧化碳浓度。j . Exp。机器人。现年49岁,443 - 452。
穆雷,湄(1997)二氧化碳和植物响应。约翰•威利和儿子,纽约。
Nemani,水银血压计基林,C.D.桥本,H。、快活、高家俊风笛手,南卡罗来纳州,塔克C.J.、Myneni R.B. &运行S.W.(2003)气候驱动增加在全球陆地净raybet雷竞技最新初级生产从1982年到1999年。科学,300,1560 - 1562。
牛顿,P.C.D.克拉克,H。贝尔,林祖嘉&格拉斯哥,E.M.(1996)交互的土壤湿度和二氧化碳升高地上生长速率、根长密度和气体交换的泥炭温带牧草。j . Exp。机器人。,771 - 779。
Norby, r & Cotrufo年报(1998)垃圾质量的问题。自然,396年,17 - 18。
抱有D。Arnone,正当第三,& Korner h(2001)原位大气中的二氧化碳升高对叶片的影响冻结阻力和碳水化合物在原生温带草原。安。机器人。,87,839844。
R.B. Olesniewicz K.S. &•托马斯(1999)的影响菌根生物量生产和固氮殖民的刺槐(洋槐pseudoacacia)幼苗生长在大气二氧化碳浓度升高。新植醇。,142,133 - 140。
Ottman, M.J.金博,文学士学位品特,P.J.墙,G.W.万德利普,R.L.莱维特,S.W.、LaMorte R.L.T.J.马提亚,公元&布鲁克斯(2001)升高的二氧化碳在干旱条件下高粱生物量增加。新植醇。,150,261 - 273。
Owensby,刚建成时私家侦探,科因、火腿、J.M.、Auen L.M.纳普A.K.(1993)生物质生产的高草草原生态系统暴露于环境和高二氧化碳。生态。达成。,644 - 653。
Palta, J.A. &诺贝尔,注:(1989)影响土壤O2和CO2为龙舌兰deserti根呼吸。杂志。工厂。,76,187 - 192。
锅,Y。所以,J.M.McGuire,公元、Kicklighter D.W.、Pitelka L.F.希巴德,K。皮尔斯,l。、跑步、S.W.、Ojima科学博士帕顿,j &丢弃科学博士(1998)模拟陆地生态系统对大气中的二氧化碳升高的反应:比较模拟植被/生态系统生物地球化学模型的建模和分析项目(VEMAP)。环境科学,114,389 - 404。
帕特森,D.T.(1986)反应的大豆(大豆)和三个C4草坪杂草二氧化碳浓缩在干旱。WeedSci。,34岁,203 - 210。
帕特森,D.T.、燧石、密纹唱片和Beyers J.L.(1984)影响的二氧化碳浓缩在C4杂草和C3作物之间的竞争。Sci的草。,32岁,101 - 105。
菲利普斯环马丁内斯,随机变数阿罗约,L。西奥多,贝克,,Killeen T。刘易斯,S.L. Malhi Y。门多萨,点尼尔D。巴尔加斯,期票Alexiades, M。Ceron C。Di凝花,。欧文,T。Jardim,。帕拉西奥斯,W。,m & Vinceti Saidlas b(2002)大llianas越来越占主导地位,亚马逊森林。 Nature, 418, 770-774.
皮尔克,R.A.Marland, G。,贝茨追逐,T.N.伊士曼,。奈尔斯,J.O.、他、科学博士和运行,S.W.(2002)土地利用变化和景观的影响对气候系统动力学:关联辐射以外的气候变化政策raybet雷竞技最新温室气体的影响。费罗斯。反式。r . Soc。Lond。,360,1705 - 1719。
住Pinelli P &洛雷托,f(2003) 12二氧化碳排放来自不同代谢途径以照亮黑暗的C3和C4叶子在低,大气二氧化碳浓度升高。j . Exp。机器人。现年54岁的1761 - 1769。
脑桥,T.L. & Welschen R.A.M.(2002)的呼吸率过高的商用桌叶室。并发症与净光合作用的测定。植物细胞包围。25, 1367-1372.
要隘,h(1998)生长缓慢的植物物种和nutrient-stressed相对强烈回应二氧化碳升高?全球变化生物学观点》4,693 - 697。
要隘,h &允许>。(2003)植物生长和竞争在二氧化碳升高:成功者、失败者和官能团。新植醇。,157,175 - 198。
要隘,h & Perez-Soba m(2001)植物的生长反应在最优条件下二氧化碳升高。环境科学,129,1 - 20。
波特凡,c & Vasseur l .(1997)长期的二氧化碳浓缩一个牧场的社区。生态学,78,666 - 677。
Rachmilevitch, S。表兄弟,A.B. &布鲁姆A.J.(2004)硝酸盐同化在植物芽取决于光呼吸。Proc。国家的。美国私立高中科学。,101,11506 - 11510。
麝香猫,聚合度Peresta g . &德雷克,人(2005)十七年的二氧化碳升高在切萨皮克湾湿地曝光:持续但对比反应的植物生长和二氧化碳吸收。全球变化生物学观点》,369 - 377。
Reddy,投资者&霍奇H.F. (eds)(2000)气候变化和全球作物生产力raybet雷竞技最新。CABI出版社,纽约。
Reekie,例如& Bazzaz碰头(1991)物候学和经济增长在四个年度物种生长在环境和高二氧化碳。可以。j .机器人。,69,2475 - 2481。
Reekie, J.Y.C.,公关& Reekie Hicklenton如(1997)的互动影响二氧化碳浓缩和昼长在佩妮矮牵牛生长和发育。安。机器人。、80、57 - 66。
帝国,P.B.球,J。,Tilman D。Craine, J。埃尔斯沃思,D。Tjoelker, M。李,T。Wedin D。、Naeem年代。Bahauddin D。Hendrey, G。何塞•S。弗拉格,K。哥特,j . & Bengston w(2001)植物多样性提高生态系统对二氧化碳和氮沉降增加的反应。自然,410,809 - 812。
卢温尼,j . &盖尔j .(1985)高水平的二氧化碳的影响在黑暗的呼吸和生长的植物。植物细胞包围。8, 623-628.
卢温尼,J。盖尔,J。& Mayer, A.M. (1993a) Photosynthesis, Respiration and dry matter growth of Lemna gibba, as affected by day/night [CO2] regimes. In: Photosynthetic Responses to the Environment (eds H.Y. Yamamoto & C.M Smith), Vol. 8, pp. 201-206. American Society of Plant Physiologists Series, Rockville, MD.
卢温尼,J。盖尔,J。& Mayer,点(1993 b)减少呼吸的高二氧化碳环境和由此产生的测量误差的呼吸与氧电极。安。机器人。,72,129131。
罗杰斯,第三世彼得森,:、McCrimmon J.N. &治愈,J.D.(1992)反应植物根系的大气二氧化碳浓度升高。植物细胞包围。15, 749-752.
罗杰斯,第三世、Runion G.B. & Krupa S.V.(1994)植物对二氧化碳浓缩强调根和根际。环绕。Pollut。,83,155 - 189。
Seneweera, S。Ghannoum o . &康罗伊,摩根大通(J.P.(2001)因素造成里生根发芽干旱的影响C4光合作用和生长的草黍coloratum二氧化碳分压升高。欧斯特。j .植物杂志。,28岁,451 - 460。
谢弗,K.V.R.奥伦,R。赖,C.-T。& Katul G.G.(2002)在一个完整的温带森林生态系统水文平衡环境和大气中二氧化碳浓度升高。全球变化生物学观点》,895 - 911。
施莱辛格,w•h•&里胥特博士j .(2001)有限公司碳储存在森林土壤和凋落物实验情节在大气中的二氧化碳增加。自然,411,466 - 469。
萨基,杰(1985)O2-insensitive光合作用C3植物。它的发生和一个可能的解释。植物杂志。,78,71 - 75。
光泽,j .(1994)反馈控制基因的表达。Photosynth。Res, 39岁,427 - 438。
自信司令部(1998)泛黄和光合衰退大麦主要叶子在大气中的二氧化碳浓缩。杂志。工厂。,103,193 - 199。
西姆斯,D.A.程,W。罗,y & Seemann jr(1999)光合适应高二氧化碳向日葵树冠。j . Exp。机器人。,645 - 653。
Sionit: &帕特森D.T.(1984)反应的C4草本植物与大气中的二氧化碳浓缩。即辐照度的影响。环境科学,65 - 34。
史密斯堡。Huxman,式样Zitzer,旧金山那里查莱特,T。N., Housman, D.C., Coleman, J.S., Fenstermaker, L.K., Seemann, J.R. & Nowak, R.S. (2000) Elevated CO2 increases productivity and invasive species success in an arid ecosystem. Nature, 408, 79-82.
副部,F.X.,h &夏基Medrano杰(1993)反馈限制光合作用的菜豆l .生长在高二氧化碳。植物细胞包围。16, 81-86.
阶梯,P。卡特尔,M。,月亮特区。罗西,。Hungate,文学士学位,g . &德雷克Hymus伞齿轮(2002)大气中的二氧化碳降低食草动物丰度升高,但叶子脱落率增加。全球变化生物学观点》,658 - 667。
施迪,m(1991)不断上升的二氧化碳水平和碳低光合细胞潜在的意义。植物细胞包围。14, 741-762.
施迪m &两部,a(1999)之间的交互高浓度二氧化碳和氮营养:生理和分子背景。植物细胞包围。22, 583-621.
圣俄梅珥、l·霍法、克里(1983)二氧化碳浓度升高和整个植物衰老。生态学,64,1311 - 1314。
竹内,Y。Kubiske,工程师Isebrands, j。、Pregtizer K.S.Hendrey, G。& Karnosky, D.F. (2001) Photosynthesis, light and nitrogen relationships in a young deciduous森林的树冠在露天下二氧化碳浓缩。植物细胞包围。24, 1257-1268.
Temperton, V.M.Grayston,中华民国杰克逊,G。巴顿,C.V.M.米勒德,p &贾维斯P.G.(2003)二氧化碳浓度升高对增长的影响和赤杨皮固氮中需长期田间试验。树杂志。,23岁,1051 - 1059。
托马斯,R.B. &应变,开国元勋之一B.R.安贝德卡对(1991)根限制因素在棉花幼苗的光合适应生长在高二氧化碳。植物杂志。,96,627 - 634。
汤普森G.B.、棕色、J.K.M. &伍德沃德F.I.(1993)主机二氧化碳的影响,氮和水的供应对小麦白粉病和蚜虫的感染。植物细胞包围。
16,687 - 702。
汤普森G.B. &德雷克伞齿轮(1994)昆虫和真菌C3莎草和C4草地上暴露于大气中的二氧化碳浓度升高在敞篷的钱伯斯。植物细胞包围。
17日,1161 - 1172。
组织,D.T.Megonigal,摩根大通(J.P. &托马斯R.B. (1997)固氮酶活动升高CO2和N2固定刺激的热带N2-fixing树。环境科学,109年,只愿降价。
批,M.T. &亚历山大,J.D.(1993)植物水关系和二氧化碳升高的影响:一个回顾和未来研究的建议。Vegetatio 104/105 47 - 62。
van Oosten j j。& Besford保留时间(1995)气体交换之间的一些关系,生物化学和分子生物学的番茄植物的光合作用在叶片发育后转移到不同的二氧化碳浓度。植物细胞包围。18, 1253-1266.
病房里,J.K. &应变,开国元勋之一B.R.安贝德卡对(1997)低和高二氧化碳分压的影响从不同海拔高度的拟南芥的生长和繁殖。植物细胞包围。,20岁,254 - 260。
病房里,J.K. &应变,开国元勋之一B.R.安贝德卡对(1999)升高的二氧化碳研究:过去、现在和未来。树杂志。,19岁,211 - 220。
沃特,jr、新闻、m &快速焊接管(2000)二氧化碳升高诱发生化和超微结构的变化C4的谷物,高粱。植物杂志。,123,1143 - 1152。
d Wedin D.A. &·蒂尔曼(1996)氮负荷和物种组成对草原的碳平衡。科学,274,1720 - 1723。
Weerakoon,波长计Olszyk, &莫斯博士(1999年)大桥街氮营养对响应的影响水稻幼苗的二氧化碳。阿格利司。Ecosyst。环绕。,72年1 - 8。
威尔逊,K.B.卡尔森,T.N. & Bunce j(1999)反馈显著影响模拟二氧化碳对季节性蒸散的影响来自两个农业物种。全球变化生物学观点》5,903 - 917。
Wullschleger金丝Ziska, L.H.& Bunce, H.A. (1994) Respiratory response of higher plants to atmospheric CO2 enrichment. Physiol. Plant. , 90, 221-229.
Zak,湄福尔摩斯,真实再现Finzi,交流陶伟洪Norby, r &施莱辛格(2003)下土壤氮循环二氧化碳升高:合成的森林面对实验。生态。达成。,1508 - 1514。
Ziska L.H.(2000)升高的二氧化碳对产量损失的影响从C3和C4杂草在野外种植的大豆。全球变化生物学观点》6,899 - 905。
Ziska L.H. (2001 a)适应增长的温度改变了温度依赖刺激的光合作用升高的二氧化碳Albutilon落叶。杂志。工厂。,1雷竞技csgo11,322 - 328。
Ziska L.H. (2001 b)变化之间的竞争力C4作物和杂草C3与二氧化碳升高。Sci的草。现年49岁,622 - 627。
Ziska L.H. (2003 a)评价六个入侵物种的生长反应过去、现在和未来的二氧化碳浓度。j . Exp。机器人。现年54岁的395 - 401。
Ziska L.H. (2003 b)产量损失评估领域高粱从C3和C4杂草随着二氧化碳。WeedSci, 51岁,914 - 918。
Ziska, L.H. & Bunce j(1993)二氧化碳和温度升高的影响种子发芽并从土壤出现。34岁的作物Res。147 - 157。
Ziska, L.H. & Bunce j(1995)增长和三个大豆品种的光合响应同步增长的温度和二氧化碳的增加。杂志。工厂。,94,575 - 584。
Ziska L.H. & Bunce j (1997 a)的影响,增加二氧化碳浓度对光合作用和生长的刺激C4作物和杂草。Photosynth。现年54岁的Res。199 - 208。
Ziska L.H. & Bunce j (1997 b)温度的作用在决定刺激的二氧化碳同化二氧化碳浓度升高的大豆幼苗。杂志。工厂。,100,126 - 132。
Ziska, L.H. & Bunce j(1998)的影响日益增长的温度和二氧化碳浓度在大豆幼苗呼吸作用和光合作用的比例。全球变化生物学观点》、4、637643。
Ziska L.H. &·考尔菲德,粮农组织(2000年)上升的二氧化碳和豚草花粉生产常见,已知allergy-inducing物种:对公共卫生的影响。欧斯特。j .植物杂志。893900年,27日。
Ziska, L.H.吉哈德,D.E.Frenz, D.A.福克纳,轮歌手,最初& Straka j(2003)城市气候变化的先兆:常见的豚草,城市化和公共卫生。raybet雷竞技最新j .过敏中国。Immunol。,1雷竞技csgo11,290 - 295。
Ziska, L.H.Ghannoum, O。贝克,j.t康罗伊,J。Bunce,正当小林,K & Okada, m(2001)地面的全球视角,评估环境的二氧化碳的反应植物大气中的二氧化碳。全球变化生物学观点》,789 - 796。
Ziska, L.H.莫里斯,严峻&戈因E.W.(2004)定量和定性评价选择小麦品种发布了自1903年以来增加大气二氧化碳:可以产生对二氧化碳的敏感性是一个因素在小麦表现?全球变化生物学观点》,1810 - 1819。
Ziska, L.H.,那么自信,司令部& Bunce j(1999)升高的影响二氧化碳的增长和气体交换三个C4物种不同二氧化碳泄漏率。杂志。工厂。,105,74 - 80。
Ziska, L.H.、自信、钢筋和克雷默,测向(1995)可逆性的光合适应瑞士甜菜甜菜和增长的二氧化碳浓度升高。杂志。工厂。,95,355 - 364。
Ziska, L.H.Weerakoon, W。Namuco,运行系统& Pamplona, R. (1996) The influence of nitrogen on the elevated CO2 response in field-grown rice. Aust. J. Plant Physiol., 23, 45-52.
继续阅读:净初级生产力
这篇文章有用吗?
推荐项目


读者的问题
-
跟不上6个月前
- 回复