光合作用
光合作用是一个相对容易理解的过程,可以用生化方程来表示。为C3植物该方程最初由Farquhar等人(1980)提出,被称为“Farquhar模型”。该模型描述的活动二磷酸核酮糖羧化酶与它的两个竞争底物二氧化碳和氧气的关系,在经典的米克利斯-门滕酶动力学方面。我们有羧化活性
其中Vcmax是最大羧化速率,C和O是二氧化碳和氧气的浓度叶绿体, r为CO2补偿点,Kc和Ko分别为羧化和氧化的米凯利斯常数。Rd是线粒体呼吸作用。
羧化作用通常受到RuBP(主要二氧化碳受体)再生速率的限制。这取决于ATP和NADP的供应,它们本身需要来自叶绿体电子传递链的电子流,而这依赖于光吸收通过叶绿体内的光系统II在此条件下,羧化速率表示为j (C - r) A = -- Rd
其中J是电子传递的潜在速率,因子4的出现是因为叶绿体电子传递链需要四个电子才能产生一个RuBP。电子流依赖于光系统II对有用光的吸收。这由一个经验曲线方程表示,其中0表示曲率(它位于0和1之间,通常约为0.7)。
其中I为经叶片吸收率和光谱质量校正后的光子通量(Lloyd et al., 1995), Jmax为可能发生的最大电子传递速率。
为Kc、Ko和r(通常假设所有C3物种的动力学常数相同)选择适当的动力学常数值,并确定它们的温度依赖性后,就可以得到实际光合作用速率为Ac和Aj的最小值。Vcmax、Jmax和Rd与叶片含氮量有关,不仅与品种有关,而且与生长条件和冠层位置有关。关于这些常数的值、温度依赖性和生理基础的讨论,读者可以参考Wullschleger(1993)和von Caemmerer(2000)。用这种方法计算Ac和Aj的一个复杂问题是,叶绿体二氧化碳浓度C依赖于气孔的开放程度。当它们关闭时,由于二氧化碳向内扩散受到限制,C的值下降。因此,C是通过实验确定的,并且已经建立了模型来表示它的行为。
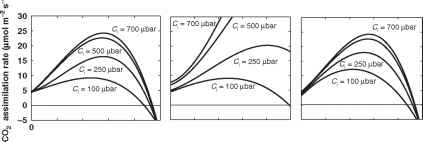
Farquhar模型的一些计算如图9.3和9.4所示。对于图9.3a所示的“典型”参数值,我们看到,在正常的CO2浓度下,光合作用速率从0°C迅速上升到20 - 30°C之间的一个相当平坦的最佳值。这与在实验室和现场实验中观察到的许多C3物种的情况非常相似。然而,当CO2增加时,最优值右移,0 ~ 20°C之间的响应变得更加剧烈。这是一个众所周知的结果,是由高的效应造成的
10 20 30 40 50 0 10 20 30 40 50 0 10 20 30 40 50
图9.3 C3植物在0-50°C温度范围内的光合作用速率,采用Farquhar模型计算,作为PAR = 1000 |imol m-2 s-1时温度和不同叶绿体CO2分压的函数。(a)净CO2同化率,(b)模拟RuBP饱和CO2同化率,(c)模拟RuBP有限同化率。根据模型,a总是b和c的最小值。
10 20 30 40 50 0 10 20 30 40 50 0 10 20 30 40 50
图9.3 C3植物在0-50°C温度范围内的光合作用速率,采用Farquhar模型计算,作为PAR = 1000 |imol m-2 s-1时温度和不同叶绿体CO2分压的函数。(a)净CO2同化率,(b)模拟RuBP饱和CO2同化率,(c)模拟RuBP有限同化率。根据模型,a总是b和c的最小值。
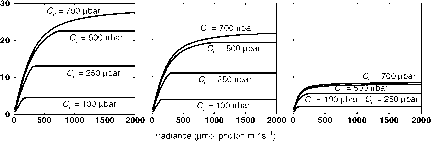
图9.4在作物(a)、热带森林(b)和针叶林(C)的四种叶绿体CO2分压Cc、25°C和200 mbar O2下,模拟CO2同化速率作为辐照度的函数。Vcmax和Jmax的值取自Wullschleger(1993),用于计算曲线:a, Vcmax - 60 |imol m-2 s-1, Jmax - 137 |imol m-2 s-1;b, Vcmax - 51 |imol m-2 s-1, Jmax - 107 |imol m-2 s-1;c, Vcmax - 25|mol m-2 s-1, Jmax - 40|mol m-2 s-1。
图9.4在作物(a)、热带森林(b)和针叶林(C)的四种叶绿体CO2分压Cc、25°C和200 mbar O2下,模拟CO2同化速率作为辐照度的函数。Vcmax和Jmax的值取自Wullschleger(1993),用于计算曲线:a, Vcmax - 60 |imol m-2 s-1, Jmax - 137 |imol m-2 s-1;b, Vcmax - 51 |imol m-2 s-1, Jmax - 107 |imol m-2 s-1;c, Vcmax - 25|mol m-2 s-1, Jmax - 40|mol m-2 s-1。
温度对rubisco羧化和加氧酶活性之间平衡的影响(von Caemmerer, 2000)。图9.3的另一个特征是,光合系统在较高的CO2浓度下趋于饱和,Wullschleger(1993)也表明了这一点,这与模拟中使用的光照水平以及参数Vcmax和Jmax的值无关(图9.4)。
9.4.2自养呼吸
呼吸作用不像光合作用那样被很好地理解。经典观点认为,自养呼吸可分为“维持呼吸”和“生长呼吸”(McCree, 1970;Thornley, 1970)。在这种模式中,假设维持呼吸释放蛋白质周转和维持内部离子浓度和梯度所需的能量,并且它也可能参与某些无效的途径和周期(Cannell & Thornley, 2000)。其值已被发现介于0.012至0.1 g C (g C)-1天-1之间,并与温度呈指数关系增长,通常被模拟为Q10为2.0。另一方面,生长呼吸取决于生长速度和正在构建的组织的化学组成,并且对温度不敏感。在模型中,这种“生长和维持”范式几乎总是被使用,尽管它可能是一种过度简化,而且它肯定忽略了物种差异。事实上,目前关于这种简单的维持呼吸模型是否适当存在争论。在数天或数周的时间内,很可能会发生一个适应过程,而维持呼吸在一定程度上依赖于基质的供应(Atkin et al., 2005;尽管存在这些不确定性,理论和实证研究表明,呼吸作用可能占光合作用的0.32-0.5整个植物,与Gifford对植物呼吸经验值的回顾一致(Cannell & Thornley, 2000a;吉福德,2003)。
9.4.3异养呼吸
异养呼吸包括来自细菌、真菌和动物的二氧化碳通量。其中,前两个在数量上是最重要的。当植物凋落物到达土壤时,它被土壤生物以实验发现的强烈依赖于温度的速率分解,在正常环境温度范围内近似于指数关系(Lloyd & Taylor, 1994;Fang & Moncrieff, 2001)。在自养呼吸的情况下,Q10通常被认为是2 (Lloyd & Taylor, 1994;Fang & Moncrieff, 2001)基于短期实验,但可能取决于长期温度。在地理范围内的长期研究中,可能适用不同的关系,因为呼吸依赖于作为微生物底物的植物材料的供应(Liski et al., 1999;Giardina & Ryan, 2000;Grace & Rayment, 2000)。“土壤呼吸”模型正在积极发展,以考虑速率的广泛差异以及温度依赖性的变化(Falloon & Smith, 2002; see also Chapter 3).
9.4.4生态系统模型
虽然可以通过实验研究叶片和整个植物的CO2通量,但在生态系统上进行有意义的实验要困难得多(见Osmond等人,2004年),因此方法通常是基于上述机制理解来模拟响应。
在过去20年里,上述方法和方程已被纳入大多数新兴的生态系统碳通量模型(Lloyd et al., 1995;Williams et al., 1996;Sellers et al., 1997),并已被用于预测碳通量将如何响应变暖和高二氧化碳。法夸尔模型用于C3光合作用只有;然而,已经开发了其他类似的模型C4植物(Collatz et al., 1992)可能占全球光合作用的五分之一(Lloyd & Farquhar, 1994)。这些模型使我们能够预测净光合作用如何随着温度升高和二氧化碳上升而变化。
虽然Farquhar模型可以很好地描述叶片的光合作用,但它在生态系统和全球模型中的应用并不简单,原因如下:
i.模型包含的参数必须与观测数据相吻合(du Pury & Farquhar, 1997),因为物种的光合成属性之间存在差异,例如,根据它们是喜欢阳光还是耐阴影,或根据树木与草本植物。此外,植物每天都要适应,而生态系统的适raybet雷竞技最新应需要更长的时间(Oechel et al., 2000),因此模型拟合可能是一个困难的过程,特别是在寒冷或干燥季节限制光合作用的压力环境中,这些原因不在这个模型中处理。
2为了控制CO2的扩散供应并使内部CO2能够计算,Farquhar模型必须与气孔行为模型耦合,其中许多巧妙的表示已经被提出(Collatz et al., 1991;环球,1995;Makela et al, 1996;Williams et al., 1996;Wang & Leuning, 1998)。所有这些都可以被描述为“经验性的”或“半经验性的”,因为它们没有机械基础,但它们仍然可以适用于观测数据。此外,空气动力学电导应该有一个子模型,因为这个术语在许多情况下限制了CO2的供应速率水分流失通过蒸腾作用和冠层表面温度(Grace et al., 1995)。
3按照设想,法夸尔模型描述了叶片层面的光合作用。虽然可以在大尺度上使用“大叶模型”(Lloyd等人,1995年),但由此可能导致的误差是不确定的(de Pury & Farquhar, 1997年),因为光合作用对光的响应是非线性的,而冠层包含完全被光照和完全或部分被遮蔽的叶子。最好将模型合并到顶棚建筑的表示中,其中表示底层(Williams et al., 1996;Wang & Leuning, 1998)。
光合作用因各种形式的胁迫而减少,最显著的是低温和缺水。生态系统模型应该在一定程度上反映相关的过程。例如,其中许多模拟了水从土壤到大气的运输,并结合了一些机制,即植物通过关闭气孔来应对干旱(Williams et al., 1996;Wang & Leuning, 1998)。
v.叶面积指数(LAI;当模型应用于生态系统尺度时,需要单位土地面积的叶片总面积)。该指数随季节和年份而变化,不太容易测量。然而,最近已有遥感产品可提供大规模的LAI估计(例如MODIS产品在edcim-swww.cr.usgs.gov / pub / imswelcome).然而,努力预测物候,冠层发育和有竞争力的相互作用来自气候数据的植物之间的差异需要与可靠和独立的现场数据进行测试(Woodward & Lomas, 2004)。这是另一个步骤,为蒸腾叶、导电边材和吸收根之间的碳的最佳分配制定规则。马格纳尼等人(2002)展示了如何使用启发式方法来实现这一目标,在分配碳以实现“最佳”解决方案之前,算法测试产生新根、茎或叶对植物的好处。
尽管有这些困难,Farquhar光合作用模型现在被用于大多数通量模型,这些模型声称是“基于机械的”。在这种情况下,我们将一个这样的模型SPA (Williams et al., 1996)应用于北方森林的数据,北方森林是最重要的生物群落之一,因为它构成了全球陆地碳总量的很大一部分,并且位于世界上预计在下个世纪升温高达8°C的地区。为此,我们对苏格兰松树的模型进行了参数化,并通过将其与芬兰Hyytiala的一个地点的通量数据拟合来调整参数值。这个网站的数据可以在互联网上免费获得。现场平均温度仅为5.4℃。该模型以小时为单位运行,对于“标准运行”,我们使用来自网站本身的数据。为了模拟变暖,我们在一年的所有季节均匀地提高温度,并对温度进行了适当的调整水蒸气压力这样空气就不会变得太干燥。降水和太阳辐射保持不变。我们还增加了叶片的二氧化碳浓度和氮浓度,以模拟这些因素在未来可能产生的影响。在该模型中,依赖温度的过程是光合作用和呼吸作用。
从图中可以看出,气候变暖对生态系统呼吸的影响最为深远,但对GPP也有显著影响(图9.5)。然而,GPP的上升幅度小于Re,因此NEP逐渐下降。在高和正常叶氮和二氧化碳条件下,当年平均温度达到10-14°C时,NEP变为负值。(图9.5)。尽管有一些特别的考虑寒冷的环境关于北方森林的研究(例如Makela等人,2004年),结果相当一般。如果我们在其他生物群落中重复这一实验,只要我们假设异养呼吸随着温度的升高而增加,总的来说,结果将不可避免地是相同的。
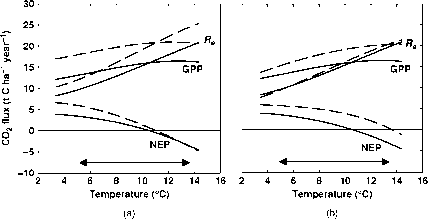
图9.5温度变化对气体交换的可能影响北方森林,使用来自芬兰Hyytiala的参数值。(a)将年GPP、NEP和Re作为正常叶氮浓度(实线)和双叶氮浓度(虚线)下温度的函数进行建模。箭头表示从当前的年平均温度(5.4°C)到考虑到100年快速变暖的可能的平均温度(13.4°)的范围。(b)与(a)相同,但CO2浓度加倍。
图9.5温度变化对北方森林气体交换的可能影响,使用来自芬兰Hyytiala的参数值。(a)将年GPP、NEP和Re作为正常叶氮浓度(实线)和双叶氮浓度(虚线)下温度的函数进行建模。箭头表示从当前的年平均温度(5.4°C)到考虑到100年快速变暖的可能的平均温度(13.4°)的范围。(b)与(a)相同,但CO2浓度加倍。
这样的模型计算使我们能够估计未来高二氧化碳世界中生物群落的碳平衡,但许多细节仍不确定。这些不确定性尤其与温度对土壤有机质分解的影响有关,如我们上文所讨论的,及其对氮和其他养分有效性的影响(如第八章所讨论);以及二氧化碳升高与极端温度的相互作用,在极端温度下,尚未建模的过程可能会发挥作用。这些过程包括在低温下,当光收集色素系统被霜冻破坏时光合作用的下调(Makela et al., 2004),以及当温度低到严重限制细胞分裂和生长时,高二氧化碳的影响(Morison & Lawlor, 1999)。
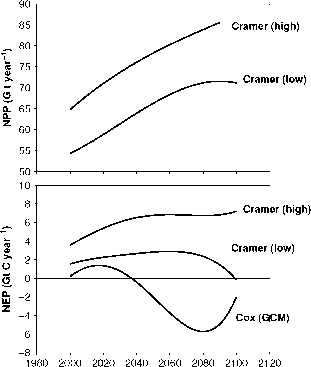
在其他模型中也可以看到与图9.5中非常相似的结果,这些模型包含了我们上面概述的假设。我们将提到两种情况。Cramer et al.(2001)比较了使用气候数据运行的六个动态植被模型的结果,根据哈德利中心气候模型的预测,这些数据代表了未来100年预计会发生的情况,假设根据IS92a情景继续排放化石燃料。raybet雷竞技最新所有的模型都表明,NPP起初是由于二氧化碳升高的施肥效应而增加的,总体来说是有益的全球变暖的影响NPP。最高和最低的模型如图9.6所示。尽管它们在总NPP上有很大差异,但增长速度是相同的,目前每十年约为2%(每十年约2.6 Pg)。另一项基于观测数据的研究显示了类似的生产力增长:Nemani等人(2003)显示,1982年至1999年,NPP每十年增长约3.5%(每十年约3.4 Pg)。
然而,Cramer等人(2001)报道的所有模型中,随着CO2饱和的发生和自养呼吸的增加,NPP的增长率下降(图9.6)。NEP在一个模型中下降得很快,但在其他模型中下降得更缓慢。在其中一个模型中,2100年全球NEP为负,此后土地成为碳源(图9.6)。
这些模型(Cramer et al., 2001)是根据使用GCM生成的气候数据运行的。raybet雷竞技最新一个更有野心的想法是将生态系统模型与GCM耦合起来,以至少获得一些反馈。例如,随着地球表面变暖,如果生态系统呼吸确实随着温度急剧增加,这将使大气中增加更多的二氧化碳,从而导致更大的变暖。这样的反馈,无论是积极的还是消极的,在气候系统的行为中都可能非常重要。raybet雷竞技最新Cox等人(2000)尝试使用一个相当简单的生态系统模型与哈德利中心气候模型HadCM3耦合(图9.6)来做到这一点。raybet雷竞技最新由于上述正反馈,变暖加速,呼吸作用如此之大,以至于生物圈成为一个巨大的来源(NEP早在2050年就变为负值,亚马逊雨林面积下降,这也是Nepstad等人从截然不同的考虑中提出的,1999年)。这是唯一一次尝试碳循环模型到气候模型,反馈不raybet雷竞技最新是那么强(Friedlingstein et al., 2003)。
这些预测是否有根据还有待观察。考克斯等人的一个批评。(2000)是异养呼吸被建模为一个简单的Q10
年
图9.6基于Cramer等(2001)植被模型的全球NPP和NEP预估。在上面的面板中给出了模型的结果,给出了NPP的最高和最低估计值。两者都在100年内增长,但其中一个达到了一个平台期。在下面的图中,全球新经济关系在小写情况下将降至零。与Cox等人(2000)完全不同的结果作为对比:在这里,NEP在今天的几十年内变为负值。
年
图9.6基于Cramer等(2001)植被模型的全球NPP和NEP预估。在上面的面板中给出了模型的结果,给出了NPP的最高和最低估计值。两者都在100年内增长,但其中一个达到了一个平台期。在下面的图中,全球新经济关系在小写情况下将降至零。与Cox等人(2000)完全不同的结果作为对比:在这里,NEP在今天的几十年内变为负值。
和温度的关系。在现实中,我们不知道这是否会在很长一段时间内发生,因为最终土壤中容易使用的碳基质会耗尽。事实上,当考察宽纬度范围内的土壤呼吸时,发现呼吸不是温度的指数函数,而更接近于线性关系(Liski et al., 1999;Giardina & Ryan, 2000;Grace & Rayment, 2000;此外,土壤碳与养分循环的相互作用对土壤肥力的形成至关重要生态系统生产力(见第八章)土壤有机质在变暖条件下的长期表现仍然是一个悬而未决的问题。
这篇文章有用吗?
读者的问题
-
梅丽莎2天前
- 回复
-
基拉19天前
- 回复
-
dawit sebhat1个月前
- 回复
-
凯瑟琳2个月前
- 回复
-
manlio输球2个月前
- 回复