光合作用
棉花的树冠生长不断升高[CO2]回应增加太阳辐射率增加的光合作用(图8.1)。
表8.1。治疗结构实验在自然阳光棉花环境室(梁单元)过去10年。
表8.1。治疗结构实验在自然阳光棉花环境室(梁单元)过去10年。
一年 |
品种 |
温度(°C)日夜 |
(|更易与摩尔1) |
评论和引用 |
外贸1 - 1989 |
DPL 50 |
15/7,20/10,25/15,30/20,35/25 |
350年,700年 |
70天出现,雨水丰沛,受精(稳压器 |
Reddy等等。,1994 a, b, 1995 a, b) |
||||
外贸2 - 1989 |
DPL 50 |
30/22 |
几个[CO2] |
短期的,几周在开花期间,水分充足 |
受精(投资者Reddy等等。1995 b) |
||||
外贸3 - 1989 |
DPL-50 |
几个高温 |
600年 |
4周在果期期间,落花的研究 |
(稳压器Reddy et al ., 1994 a, b, 1995) |
||||
外贸4 - 1990 |
Pima-S-6 |
20/12,25/17,30/22,35/27,40/32 |
350年,700年 |
64天出现,雨水丰沛,受精(投资者 |
Reddy等等。,1995 a, b) |
||||
外贸5 - 1990 |
Pima-S-6 |
25/17,30/22,35/27 |
350年,700年 |
开花到成熟,雨水丰沛,受精(投资者 |
Reddy等等。,1995 a, b, c) |
||||
外贸6 - 1990 |
Pima-S-6 |
四个高温 |
700年 |
开花季节,水果(境Reddy保留研究 |
等等。,1997;投资者Reddy等等。,1993) |
||||
外贸7 - 1991 |
Pima-S-6 |
30/22 |
350年,450700年 |
95天出现,三个干旱压力水平(投资者 |
Reddy和霍奇斯,1998) |
||||
外贸8 - 1991 |
DES 119 |
30/22 |
350年 |
四个氮水平(投资者Reddy等等。,1997 b;稳压器Reddy |
等等。,1997) |
||||
外贸9 - 1992 |
DPL 5415 |
26/18,31/23,36/18 |
350、450、700 |
从出现60天,雨水丰沛,受精(境 |
Reddy等等。,1998)
Reddy等等。,1998)
外贸10 - 1992 DPL 5415 26/18, 31/23, 36/18 350450700人
外贸1 1 - 1993 DES 19 350700 30/22
外贸12 19 - 1993 DES 1 30/22 350450700人
外贸13 - 1994 AcalaMaxxa 20/1, 25/1, 30/22, 35/27, 40/32 360720 HS-26 DPL 51
外贸14 - 1994 DPL 51温度:长期女士360年7月,700年
出口温度15 - 1995 DPL 51: 1 995环境,1 995 360,720
ambient-2, 1995环境+ 2、5和7
外贸16 - 1996 NuCot33 30/22 360720人
外贸17 - 1996 NuCott33 26/26 360
外贸18 - 1997 Nucot33 30/22 360720人
外贸19 - 1997 NuCot33几个短期温度[co2]年代治疗
开花到成熟,水分和受精。
从出现49天,5 N水平(投资者Reddy ef /。1997 b)
80天的出现,三个干旱压力水平(投资者Reddy ef /。1997 c)
从出现46天,水分和受精(投资者Reddy ef /。1997 d)
4周、开花期、浇灌和受精(投资者Reddy ef /, 1997)
瑞,雨水丰沛,受精(投资者Reddy ef /。,1997;1998)。
84天,5 K的水平
手动de-leafing和de-fruiting研究中,水分和受精
水缺乏的一些研究
短期的,几天,几周,雨水丰沛,受精- * - *
一年 |
品种 |
水的供应 |
营养供应 |
[CO2](更易mol-1) |
评论和引用 |
敞篷的室实验 |
|||||
1983年 |
DPL 70 |
有实力的 |
Well-fertilized |
环境、500、650 |
Ambient-no-chamber治疗还包括(金博等等。,1992 b;金伯尔Mauney, 1993) |
1984年 |
DPL 61 |
有实力的Drought-stressed |
Well-fertilized |
环境、500、650 |
Ambient-no-chamber治疗还包括(金博等等。,1992 b;金伯尔Mauney, 1993) |
1985年 |
DPL 61 |
有实力的Drought-stressed |
Well-fertilized |
环境、500、650 |
Ambient-no-chamber治疗还包括(金博等等。,1992 b;金伯尔Mauney, 1993) |
1986年 |
DPL 61 |
有实力的Drought-stressed |
高氮低氮吸收 |
环境,650 |
金伯尔等等。(1992 b);金伯尔和Mauney (1993) |
1987年 |
DPL 61 |
有实力的Drought-stressed |
高氮低氮吸收 |
环境,650 |
大麦N-removal作物种植在1986 - 1987年冬季(金博等等。,1992 b;金伯尔Mauney, 1993) |
自由二氧化碳富集(脸)实验
1989 DPL 77浇灌Well-fertilized环境,550年
1990 DPL 77浇灌Well-fertilized环境,550年
Drought-stressed
1991 DPL 77浇灌Well-fertilized环境,550年
Drought-stressed
Hendrey (1993);Dugas和品特(1994)Hendrey (1993);Dugas和品特(1994)
Hendrey (1993);Dugas和品特(1994)
0 500 1000 1500 2000
光合光量子通量密度(^摩尔rrr2年代~ 1)
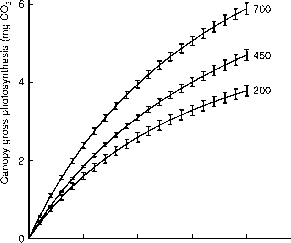
图8.1。二氧化碳和太阳辐射的影响(光合光量子通量密度,PPFD)棉花树冠总光合作用(Pg) 60 DAE晶石实验。Pg =净光合作用和呼吸作用。(投资者Reddy et al ., 1995 c)。
0 500 1000 1500 2000
光合光量子通量密度(^摩尔rrr2年代~ 1)
图8.1。二氧化碳和太阳辐射的影响(光合光量子通量密度,PPFD)棉花树冠总光合作用(Pg) 60 DAE晶石实验。Pg =净光合作用和呼吸作用。(投资者Reddy et al ., 1995 c)。
最初的斜坡上的光响应曲线和渐近线在升高[CO2]。这表明近最优生长条件,额外[CO2]导致较高的光合作用由于羧化酶固定利率的效率(更高的利率,光限制)和气体的扩散到固定地点(更高的利率,二氧化碳是主要的限制因素)。这一点上进一步说明了photo-synthetic (Pnet)反应的棉花的树冠二氧化碳和辐射在不同空气温度(图8.2)。这个实验是由植物生长环境和twice-ambient [CO2] 5点温度。美国1995年的温度在密西西比州,被用作参考,与其他温度1995 - 2°C,和1995 + 2,5和7°C。每日和季节性变化和振幅是维护。净光合作用在高和低的温度低于最优。空气温度和反应的趋势[CO2]被投资者类似报道Reddy et al . (1995 c)。三个较低的温度和环境的二氧化碳,Pnet应对辐射的增加随着温度的增加而减少。这显然反映了更高的呼吸由高温引起的植物的树冠。 The opposite effect was found with high CO2, but this was w
1995环境- 2°C •> |
•720 pmol co2摩尔360 o pmol co2摩尔”1 |
1995年环境温度 |
1995环境+ 5°C ,“ |
1995环境+ 2°C »v••»9 |
1995环境+ 7°C |
0 500 1000 1500 2000 0 500
1000年
1500 2000
0 500 1000 1500 2000 0 500
1000年
1500 2000
图8.2。太阳辐射的影响(PPFD), [CO2]和温度对棉花树冠净光合作用(Pn) 1995年晶石实验。植物从出现在各自[CO2]和温度环境。数据代表80天之后出现的树冠被拦截大约95 - 98%的入射太阳辐射。
更大的增长的结果升高[CO2]种植植物和更多年轻的叶子导致更高的光合速率。
1995 + 5 + 7°C治疗温度,植物的photo-synthetic利率在760年更易与二氧化碳mol-1环境也高。这些植物微创吐絮期开花期后不久。棉花是一种多年生植物,作为年度商业棉花生产管理。棉花植物通常设置他们的第一个水果在果期核电站附近的底部分支节点(5 - 8)(投资者Reddy et al ., 1997 b)。在最佳条件下,新果期网站产生下一个更高的主节点3天后,和在果期相同分支的下一个节点6天后。棉花植物逐步添加吐絮期,直到可用光合供应将不再支持额外的水果。作为支持高人口的营养需求吐絮期的植物增加,营养生长的营养不再可用。这导致减速并最终停止茎和叶的生长。减少茎增长反映在缓慢扩张的节间和产生更少的节点。节点产生,果期分支越来越少,因此更少的网站更多的水果。 Even under good conditions, nutrient requirements decline as bolls mature and vegetative growth resumes. In high-temperature environments (above 32°C), fruits were abscising 3-5 days after anthesis, thus voiding typical nutrient sinks and therefore promoting luxuriant vegetative growth. The high photosynthetic rates in these high-temperature environments (1995 plus 5 or plus 7°C) resulted from the relatively young population of leaves continuously being added to the top of the canopy.
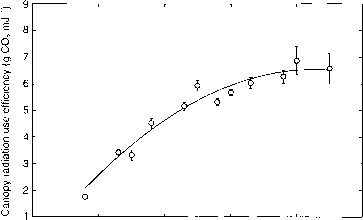
[CO2]对光合效率的影响见图8.3。通过从这一数字插值,可以确定棉花植物修复在4.3 g m - 2 MJ-1二氧化碳CO2环境(360 |更易mol-1)和6.3克二氧化碳的m - 2 MJ-1 twice-ambient [CO2]。这是造成光合效率增加25%翻倍[CO2]。这两个值代表有环境中高效的固定利率。低的光合效率是有限的可用的二氧化碳(CO2)。二氧化碳固定的效率增加[CO2]增加,但以递减的速度。今天环境(二氧化碳)的两倍,基本上所有的可行的获得在光合作用完成。
在图8.4中提供的信息显示了树冠光合作用的变化与归一化辐射整个赛季的1200 |更易与m - 2 s - 1,和自然不同的温度。这些数据说明光合速率随着时间的太阳辐射归一化,这样在辐射季节性变化不是一个混杂因素。光合作用增加,直到出现大约80天后,然后减少在整个生长季节。农作物生长环境[CO2]他们的光合速率下降超过作物生长在twice-ambient [CO2]。这是由于年龄增加light-intercepting的树冠。在高二氧化碳环境中,光合作用仍继续在一个非常健康的
200 400 600 800 1000 0
二氧化碳浓度(irmol co2 mol-1)
图8.3。大气的影响[CO2]棉花的树冠的辐射利用效率。(投资者Reddy et al, 1997 c。)
200 400 600 800 1000 0
二氧化碳浓度(irmol co2 mol-1)
图8.3。大气的影响[CO2]棉花的树冠的辐射利用效率。(投资者Reddy et al, 1997 c。)
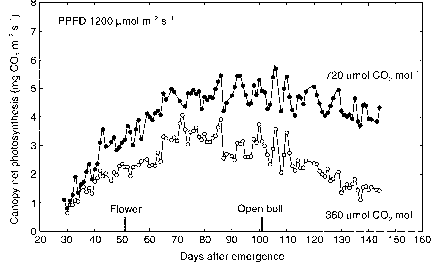
图8.4。净光合作用的时间趋势(Pn)的棉花的树冠生长在1995环境温度在360年和720年| |摩尔二氧化碳mol-1在密西西比州密西西比。光合作用连续测量,总结每隔900年代,和归一化1200 |更易与m - 2 s - 1每天整个赛季。时间第一花和成熟·鲍尔表示。(投资者Reddy et al ., 1998 b)。
图8.4。净光合作用的时间趋势(Pn)的棉花的树冠生长在1995环境温度在360年和720年| |摩尔二氧化碳mol-1在密西西比州密西西比。光合作用连续测量,总结每隔900年代,和归一化1200 |更易与m - 2 s - 1每天整个赛季。时间第一花和成熟·鲍尔表示。(投资者Reddy et al ., 1998 b)。
速度甚至在本赛季结束后,提供足够的减少碳排放来支持一些新的叶子生长在果期期间。
的平均光合速率的树冠在环境和twice-ambient [CO2]概括为利率720/360 | |摩尔二氧化碳mol-1,商议与平均季节性的温度(图8.5)。季节平均光合速率在720 |更易与二氧化碳mol-1是大约140%的360 |更易mol-1 20°C和32°C,但响应增加超过180%,接近最佳温度(代谢途径°C)。图8.6提供了棉花的相对光合反应的树冠生长在高架[CO2]与作物环境[CO2],但在一系列温度和缺水条件。本图中的数据代表光合速率比马和陆地棉的树冠上,虽然个人数据点之间存在很大差距,回归线表明,光合作用在720 | mol-1更易与二氧化碳的156%,在360年| mol-1更易与二氧化碳。光合反应twice-ambient [CO2]似乎线性即使在高。
这些结果从受控环境室通常是符合观测的影响升高[CO2] field-grown植物的光合作用。雷丁et al。(1987)报道,二氧化碳浓缩到650 | 1985敞篷的摩尔二氧化碳mol-1室实验(表8.2)雨水丰沛的棉花叶片光合作用增加超过70%。Hileman) et al。(1994)发现,叶子和树冠在富水条件下光合率[CO2]时增加了约27%
-

- 直流

1.0 - 1-1-1-1-1-1-1 -
温度(°C)
图8.5。效应的相对反应温度对棉花光合作用树冠加倍环境[CO2]。温度和光合率平均在本赛季从95%的辐射截获的树冠,直到第一次打开·鲍尔。
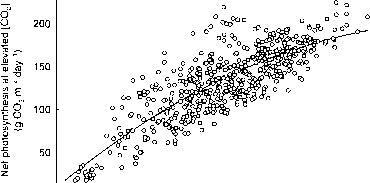
0 20 40 60 80 100 120 140 160 180净光合作用在环境[co2] (g二氧化碳rrr2普通人每天)
图8.6。日常关系总净林冠棉花光合作用植物生长环境(360 |更易mol-1)和高[CO2]。数据从五个温度的陆地棉和三个水分亏缺治疗治疗比马棉收集几天在果期。
浓缩到550 |更易mol-1在1989年面对实验(表8.2),和树冠利率在1990年增加了31%。
棉花植物生长在缺水条件下在受控环境室的光合速率下降中午叶水减少无论[CO2](表8.3;投资者Reddy et al ., 1997 c)。植物生长在720 |更易与二氧化碳mol-1固定平均超过48%的二氧化碳
表8.3。参数方程回归几个发育事件/和增长率的棉花生长在高架(700或720 |更易与二氧化碳mol-1)大气中的二氧化碳(y)作为植物生长在环境的函数(350或360 |更易与二氧化碳mol-1)二氧化碳(x) (y = b0 + b1x - b2x2)。获得的数据从植物种植在一系列温度和营养和水缺乏的条件。
表8.3。参数方程回归几个发育事件/和增长率的棉花生长在高架(700或720 |更易与二氧化碳mol-1)大气中的二氧化碳(y)作为植物生长在环境的函数(350或360 |更易与二氧化碳mol-1)二氧化碳(x) (y = b0 + b1x - b2x2)。获得的数据从植物种植在一系列温度和营养和水缺乏的条件。
回归参数 |
||||
参数 |
薄 |
b1 |
b的2 |
r2 |
发育事件 |
||||
出现广场(天) |
-0.595 |
1.016 |
- - - - - - |
0.99 |
广场花(天) |
-1.184 |
0.944 |
- - - - - - |
0.92 |
花开放·鲍尔(天) |
-1.615 |
0.996 |
- - - - - - |
0.99 |
主节点(没有。plant 1) |
0.313 |
0.998 |
- - - - - - |
0.98 |
果期(没有网站。plant 1) |
-0.999 |
1.384 |
- - - - - - |
0.95 |
(没有根。m - 2普通人每天) |
-2.460 |
1.160 |
- - - - - - |
0.94 |
(没有气孔密度。mm-2) |
-9.365 |
0.954 |
- - - - - - |
0.99 |
增长和其他进程株高(厘米Plant 1)分支长度(m Plant 1)叶面积(平方米Plant 1)植物总重量(g Plant 1)杆重量(g Plant 1)叶重(g Plant 1)根重量(g Plant 1)水果重量(g Plant 1)净光合作用 |
2.419 -103.88 -0.022 2.352 -0.0427 1.936 0.156 0.216 -107.129 |
1.026 1.168 1.248 1.324 1.434 1.240 1.298 1.256 9.012 |
-0.167 |
0.93 0.96 0.98 0.88 0.97 0.97 0.89 0.98 0.94 |
(g m - 2普通人)蒸腾 |
2.7799 |
0.923 |
- - - - - - |
0.83 |
(公斤水m - 2地面面积普通人)蒸腾 |
36.741 |
0.510 |
- - - - - - |
0.99 |
(毫克H2O m - 2叶面积s - 1) 1电弧炉N (%) |
-0.46 |
0.954 |
- - - - - - |
0.95 |
植物生长在360 |更易mol-1在一些缺水的条件。中午叶水势变化从大约-1.2 MPa到-3.0 MPa。没有明显的光合响应之间的交互植物生长在高架[CO2]和缺水条件。植物生长在环境和twice-ambient [CO2]光合作用降低了约43%作为他们中午从-1.2 MPa叶水势降低到-3.0 MPa。场结果相似。雷丁et al。(1987)测量净增加52%叶光合作用[CO2]在650 |摩尔二氧化碳mol-1 drought-stressed条件下在1995敞篷的室实验(表8.2)。Hileman) et al。(1994)在树冠光合作用增加了21%浓缩到550 | 1990面临干旱胁迫下更易与二氧化碳mol-1实验。
棉花植物应对缺N在受控环境室有点类似于植物应对水赤字(表
8.3;投资者Reddy et al ., 1997 b, c)。光合作用下降,叶片氮降低了。植物生长在twice-ambient [CO2]他们的光合作用降低了关于N-deficient条件相同的植物生长在环境[CO2]。植物种植以2%叶N在360年和720年|更易与二氧化碳mol-1 38%植物光合率低于4%叶N,和植物生长在twice-ambient [CO2]只有固定的41%的二氧化碳比植物生长在环境[CO2]。Canopy-level光合作用是植物生长在更大更高的N与环境和升高[CO2]。光合作用下降迅速在果期期间当树冠N不足,但如果N是足够的(叶N大约3%),树冠光合率保持50%更高的利率twice-ambient比环境[CO2] [CO2](投资者Reddy H.F.霍奇斯,未公开的数据)。棉花的树冠保持90%最大光合速率较低的叶片在高N - [CO2]环境比环境[CO2]。发现了类似的涨幅N的利用效率水稻(P.J.康罗伊、新南威尔士、澳大利亚、个人通信,1999)。这些结果表明,更高的收益率可能获得高——[CO2]环境随着N施肥。Field-grown棉暴露于650 |更易与二氧化碳mol-1各位叶净光合率增加了45 - 70%,根据年,没有明显的影响N的压力反应(表8.2;金伯尔et al ., 1986, 1987)。
缺钾引起的9.1%和6.8%下降为叶光合率K在环境从4%下降到2%,twice-ambient [CO2]环境,分别(投资者Reddy et al ., 1997)。然而,光合作用下降更快为叶K从2%下降到1%。叶光合作用降低了10.6%和15.7%的1% K植物比叶2% K植物在环境和twice-ambient [CO2],分别。钾营养和[CO2]没有交互影响光合作用叶片K浓度高于2%时,但光合作用下降到接近零在植物生长极低K .因此,看来压力是否引起的水赤字或N和K不足,很少或没有交互[CO2]对光合作用的影响。巴雷特和吉福德(1995)发现了类似的结果与P营养和[CO2]浓缩。
这篇文章有用吗?