种子发芽
热带植物的种子雨林的树木显示一个较大的种间发芽时间范围。在马来西亚森林中330个物种的样本中,65%的物种在浅阴影苗圃中播种新鲜种子后的20周内萌发(Ng 1980)。35%的种子需要超过20周的时间才能发芽,种子周围通常有坚硬而厚的种皮或内皮。对萌发生理学的研究表明,许多因素可以导致热带雨林树木种子萌发延迟(Vazquez-Yanes & Orozco-Segovia 1993)。这些因素包括成熟时种子含水量低、种皮坚硬、胚体积小、发育早期以及存在化学发芽抑制剂。发芽快的种子成熟时种子含水量高,种皮柔软。许多这样的物种很难储存。如果干燥,它们就失去活力,如果储存在高含水量的环境中,它们就会发芽。这使得种子生理学家开发了一种基于种子存储潜力的分类系统。正统种子(Roberts 1973)是指那些可以在休眠状态下储存较长时间(许多月)的种子,通常种子含水量相对较低。 Recalcitrant seeds are the rapid germinators that cannot withstand drying. Many tropical tree species produce large, soft-coated seeds of high water content that are recalcitrant.
更正统的物种的种子休眠可以通过许多方式来实现。坚硬和不透水的种皮会从物理上阻止萌发,胚在萌发前可能需要发育,或者环境条件可能会诱导休眠。具有硬皮或持久内皮的种子在发芽前可能需要动物或环境因素的划痕。高温可能是另一个因素。巴尔沙的小种子,锥体,保持休眠,因为一个不渗透的种皮。浅表火灾的热量或与大间隙小气候相关的温度波动可导致种皮破裂,使萌发得以进行。raybet雷竞技最新阴影似乎是最重要的诱导休眠的种子热带树木。光母细胞种子对撞击辐射的光谱组成有反应。人们普遍认为光敏色素是介导种子光敏作用的化学物质(Vazquez-Yanes & Orozco-Segovia 1996a)。
-
 ratio gradient. Bars represent ±1 SD. After Vazquez-Yanes et al. (1990).")
- 图5.4钝叶天眼(Cecropia obtusifolia)沿红:远红(R/FR)比值梯度萌发。条形代表±1sd。源自Vazquez-Yanes等人(1990)。
 ratio gradient. Bars represent ±1 SD. After Vazquez-Yanes et al. (1990).")
波长约为660 nm的红光会刺激光敏色素的生理活性形式的形成,从而引发种子的萌发。约730 nm波段的远红光启动活性光敏色素向非活性光敏色素的转换,并引起休眠。红光波段和远红光波段的相对通量率(R: FR)是光母细胞种子休眠或萌发的关键光谱质量触发因子。钝叶天眼种子对R: FR变化的萌发响应如图5.4所示。森林中不同微点的R: FR有很大差异。树冠的叶子吸收红光,使森林下层相对丰富的远红波长。因此,森林遮荫诱导光母细胞种子休眠,但冠层间隙可触发萌发。凋落叶和土壤也有选择性的光谱吸收率,倾向于降低通过它们的光的R: FR。太阳斑点,即短暂的阳光直接照射,在地球上的大部分地点不规则地出现森林地面。由于它们的高R: FR可能足以引发萌发,但由于森林下层阴影光的低R: FR重新建立而产生的光还原可能会在萌发发生之前重新实现休眠。具有光母细胞种子的热带树木对森林中R: FR自然范围的萌发反应在种间,有时在种内表现出相当大的差异(Vazquez-Yanes & Orozco-Segovia 1996a;梅特卡夫1996)。光可能不是促使休眠种子萌发的唯一因素。有证据表明,在某些物种中,光与温度相互作用。例如,Orozco-Segovia等人(1987)发现,Urera caracasana的种子需要暴露在25°C的白光下4小时才能获得较高的发芽率,但在35°C下,光照30分钟就足以达到相当的发芽率。
潜在地,环境诱导的休眠可以让种子在雨中保持活力森林土很长一段时间。来自墨西哥的几个“先锋”物种的种子在实验室黑暗中储存时,完全吸收的种子保持了5年以上的高活力(图5.5)。paniculatus的种子在马来西亚雨林的网袋中埋藏3年后,没有显示出活力的下降(Kanzaki et al. 1997)。有证据表明,种子对发芽环境的要求可能随着时间的推移而变化
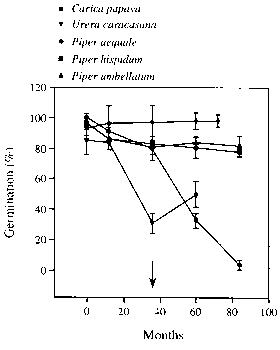
图5.5全吸收种子在黑暗中不同贮藏期后的萌发情况。3年后,同一种属的干种子发芽率均接近于零。箭头表示所有品种的种子在室温下干燥储存时失去所有萌发性。SD用竖条表示。源自Vazquez-Yanes & Orozco-Segovia (1996b)。
图5.5全吸收种子在黑暗中不同贮藏期后的萌发情况。3年后,同一种属的干种子发芽率均接近于零。箭头表示所有品种的种子在室温下干燥储存时失去所有萌发性。SD用竖条表示。源自Vazquez-Yanes & Orozco-Segovia (1996b)。
埋在土壤中(Vazquez-Yanes & Orozco-Segovia 1996a),在以前保持休眠的条件下更容易发芽。
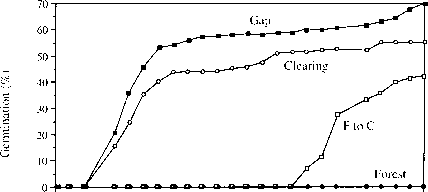
一些物种能够在阴凉处休眠,只在阳光直射下发芽,这被认为是一组生态相似的热带树种的共同特征。Swaine和Whitmore(1988)用这个作为定义“先锋”物种的关键特征。这些是快速增长的,shade-intolerant通常只在森林的间隙或其他早期演替地点发现的物种。越那些在物种的萌发通常不依赖于荫蔽的程度。Swaine & white -more(1988)将这些物种称为“非先驱者”。很少有人尝试检验Swaine & white -more提出的二分法的有效性。Raich & Gong(1990)调查了43种马来西亚树种在全日照PAR含量为60%、40%和1.2%的空地、林隙和林下层的萌发情况。只有7种树种在所有地点的萌发情况相同。在开阔地中,有16种种子萌发率低,或完全失败;有22种种子在林下萌发率高于林下,有12种种子在林下萌发率高于林下。有些树种在林下层很少发芽,但当这些种子转移到空地上时就很容易发芽(图5.6)。清理条件杀死了许多物种的种子。Raich & Gong(1990)认为,在萌发反应上,物种的明确两分法是不容易辨别的。种子对环境的反应更为复杂。 This was also the picture to emerge from a study of 19 species from tropical West Africa (Kyereh et al. 1999). Only three species germinated in significantly lower proportions in complete darkness than a light treatment and only one species was found to have a germination response to variation in R : FR. All the species showed some germination in forest understorey conditions. The sample included several species that are strongly light-demanding such as Ceibapentandra and Ricinodendron heudelotii.
种子周围的厚壳不一定会延迟发芽。在Mezzettia parviflora中,种子有3-4毫米厚的木质覆盖物,来自于中间的被皮(Lucas et al. 1991b)。这可以保护大型种子免受大多数攻击者的攻击。猩猩可以用牙齿咬开一些种子。然而,Mezzettia的幼苗可以很快地从种子中发芽。这是因为外壳周围有一层特殊的易碎短囊藻(石细胞),一端的壁上有一个小塞子。这为壳提供了一个内置的弱点,允许种子的膨胀压力打开它。哺乳动物的食种动物就无法借助这种设计,因为插头和带子太窄,它们的牙齿无法利用。吃种子的甲虫是如此之小,以至于它们的活动规模比壳的硬度更重要。弱带的硬度与壳的其他部分没有明显的差别,所以捕食者不能利用它。
platanifolia的大种子含有大量的粘液(27%)(Garwood 1985)。黏液吸水非常快(10分钟7g水~果实)。这可能有助于种子发芽和幼苗在巴拿马巴罗科罗拉多岛季节性干燥的森林中生长。
推迟发芽
为什么不同物种之间的发芽时间有这么大的差异?等待的主要好处是增加二次传播的可能性,并可能使种子的发芽错开,使一些种子逃脱不利时期,如干旱,它们本可以在干旱时期成为幼苗。等待的缺点是种子的死亡率。快速发芽者通过迅速长成幼苗来躲避种子捕食者,尽管一些种子捕食者也会攻击幼苗的子叶。快速的种子萌发有助于捕食者的饱足,因为这意味着捕食者利用种子的时间更少。具有检测环境质量的机制并相应地发芽或保持休眠的种子具有延迟萌发和快速萌发两种选择的好处。
发芽时间
巴拿马的巴罗科罗拉多岛每年都有相当明显的旱季。热带雨林中的物种在萌发时间方面表现出三种主要的气候季节性模式(Garwood 1983): 42%的物种在旱季传播种子,并保持休眠,直到降雨开始和萌发发生;40%的品种在湿润季节分散发芽;18%的物种在一个雨季就已经散去,但直到下一个雨季才开始发芽。
发芽和凋落物
如上所述,森林叶凋落物可作为选择性滤光器,透射还原R: FR的光谱组成,从而影响种子的萌发。凋落物也可以作为幼苗建立的物理障碍。小的幼苗可能无法将树叶推开,并会在凋落物下的弱光条件下死亡。对于三种树种,它们之间有直接的相关性种子大小以及在实验室通过凋落物萌发成功(Vazquez-Yanes & Orozco-Segovia 1992)。在荫凉条件下,凋落物层对“不耐荫凉”物种种子萌发和发芽的负面影响大于“耐荫凉”物种(Molofsky & Augspurger 1992)。然而,在“不耐阴影”的物种中,对凋落物抑制的敏感性有相当大的范围。在强光条件下,垃圾实际上对一些物种有益,比如古斯塔维亚。种子可能比没有凋落物时更凉爽更潮湿。空隙中的垃圾可能有利于更耐阴的物种的殖民。在波多黎各的森林中,研究发现清除凋落物可以显著增加幼苗密度(Guzman-Grajales & Walker 1991)。增加的主要来自小种子物种,如天眼草(Cecropia schreberiana)和Chionanthus domingensis。大种子物种要么不受凋落物处理的影响,要么在凋落物清除地点变得稀少。种子大小对萌发和幼苗建立的影响已被进一步的实验证实(Everham et al. 1996)。 Litter removal and disturbance of the soil surface (scarification) have been reported as increasing the density of germinants by a factor of 2.5 in gaps in lowland dipterocarp forest in Danum Valley, Sabah, Malaysia
-我-,,,我,,”,我,我”。我-.-i-i-i-i-i-i-i-i -
0 5 10 15 20 25
-我-,,,我,,”,我,我”。我-.-i-i-i-i-i-i-i-i -
0 5 10 15 20 25
播种后数周
图5.6毛trema tomentosa (ull -maceae)在林下、林隙和空地上的累积发芽时间。第15周,将一盆种子(F到C)从森林底层移到空地上;第二盆留在底层。所有的盆中都有150颗种子。请注意,y轴延伸到0以下,清楚地表明在森林下层没有萌发。连续28周记录种子萌发,25周后未见萌发。各处理间最终萌发量差异显著。源自Raich & Gong(1990)。
(Kennedy & Swaine 1992)。这些种子只占土壤最上层(0-5厘米)种子库的一小部分,即使有切口,也不超过5%。在新加坡的森林中也得到了类似的结果(Metcalfe & Turner 1998)。
继续阅读:幼苗死亡率
这篇文章有用吗?
读者的问题
-
Cataldo马里诺一年前
- 回复
-
Scudamor一年前
- 回复
-
西米一年前
- 回复