幼苗死亡率
热带雨林中树苗的死亡率普遍较高,特别是新萌发的树苗。许多苗木种群动态调查很可能会遗漏早期和最严重的死亡率,因为监测微小的幼苗并正确识别它们是非常困难的(Kennedy & Swaine 1992)。一群种子的数量通常呈对数线性下降,至少在最初是这样。也就是说,在对数尺度上绘制幼苗数与线性绘制时间的关系,可以得到一条负梯度的直线(图5.9)。Li等人(1996)在哥斯达黎加的La Selva,从森林中萌发高峰后2-3个月开始,对每年的幼苗进行了跟踪研究。多物种队列的平均半衰期为2.49个月。在潮湿时期建立的队列存活率最高。个体物种的队列半衰期从0.37至40.7个月不等。像Cecropia obtusifolia和C. insignis这样的物种的半衰期非常短。像苏格拉底durissima和Welfia georgii这样的大种子棕榈树是最持久的幼苗。 There was a negative relation between seedling recruitment and seedling persistence across the species (Fig. 5.10). In other words, there was no species that was very abundant as a new germinant that had a high seedling survival. Survivorship appears to improve as seedlings get larger. Among three common tree species on Barro Colorado Island, Panama, seedlings greater than 50 cm tall showed annual survival of more than 80% (De Steven 1994).
小秧苗上森林地面都有可能是死因其中包括以下内容。
树叶、细枝、树枝和树干如雨点般不断地落在森林地面上。小苗易受损害,这是常有的事
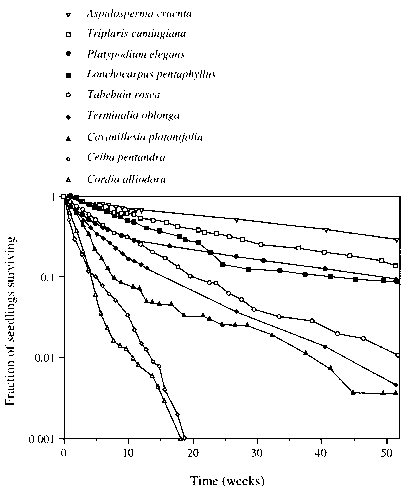
图5.9巴拿马巴罗科罗拉多岛阴影条件下9个物种的所有标记幼苗存活一年的比例。这个分数是基于在远离亲本的所有距离上存活的幼苗的总和。在距离亲本的每个距离间隔上观察到相同数量的幼苗。源自Augspurger(1984)。
图5.9巴拿马巴罗科罗拉多岛阴影条件下9个物种的所有标记幼苗存活一年的比例。这个分数是基于在远离亲本的所有距离上存活的幼苗的总和。在距离亲本的每个距离间隔上观察到相同数量的幼苗。源自Augspurger(1984)。
-
 at La Selva, Costa Rica. Longer half-lives are associated with lower seedling recruitment over the 18month period. After Li et al. (1996).")
- 图5.10在哥斯达黎加的La Selva, 75个最丰富的物种(5个或更多发芽的幼苗)的幼苗丰度与存活率之间的关系。半衰期较长与18个月期间的幼苗招募较低有关。源自Li et al.(1996)。
 at La Selva, Costa Rica. Longer half-lives are associated with lower seedling recruitment over the 18month period. After Li et al. (1996).")
致命的,从许多这些下落的物体(Aide 1987)。此外,大型哺乳动物(大象、鹿、猪、人类)在移动时会破坏森林地面的植被。幼苗长得大的一个主要好处是减少了被掉落的垃圾或动物活动破坏的风险。Clark & Clark(1989)使用塑料吸管和金属丝制成的人工幼苗来评估La Selva森林中的幼苗损害率。在一年的时间里,82%的幼苗被打翻、夷平或连根拔起。在可识别破坏因素的49%案例中,大约一半是由于掉落的碎片,另一半是由于脊椎动物的踩踏和连根拔起。Guariguata(1998)在巴罗科罗拉多岛研究了四种直立、无分枝建筑的1-2.5米高的树苗。每年约有3%的树苗受到损害。在四年的时间里,由于树苗被钉在森林地面上而导致的死亡率几乎是由于树苗断裂而失去上半部分而导致的死亡率的两倍。在研究的树种中,最具弹性的黑桤木(Alseis blackiana)能够在弯曲的树苗上产生不定根。 Interestingly, it was the most slow-growing of the four species and therefore would have the longest average residence time in the understorey.
树苗可能需要比后期更坚固的设计,以承受掉落的碎片;这一想法与从高度-直径关系的异速生长分析中计算出的安全系数的研究相吻合(King 1996;Claussen &
Maycock 1995;Rich et al. 1986)。然而,这种设计安全的方法可能存在缺陷,因为树苗倾向于在叶片上投入大量资金,使树冠质量占总质量的很大比例。因此,树苗可能并不像树干异速生长看起来那么安全;King(1987)采用了不同的方法,似乎证实了这一点。他在哥斯达黎加的La Selva用重物装载树冠物种的树冠和树苗。实验结果得出的结论是,植物的茎直径仅为最小直径的1.3-1.4倍,以防止在荫蔽下层中生长时不稳定。森林里没有强风,可能会有如此狭窄的安全限制。
幼苗捕食者
一系列不同的动物会攻击树苗,主要是为了食物,并导致它们死亡。在最初的阶段,以种子为食的动物可能会吃掉子叶,并抢夺种子的能量储备。这样的活动也会导致幼苗被拔起。草食哺乳动物可能会在幼苗变成木质之前吃掉整个幼苗。在马来西亚的帕索森林,筑巢的野猪是大型幼苗和树苗死亡的主要原因。猪用细茎建造了一个庇护所,在那里养育它们的幼崽,并为此目的从下层林下收集许多树苗。据观察,哺乳动物的食草性是Barro Colorado岛上的Virola nobilis幼苗直接死亡的主要原因,它也导致了干旱导致的死亡,因为脱叶植物无法长出足够大的根系来应对缺水(Howe 1990)。将哺乳动物排除在森林幼苗之外可降低死亡率(Osunkoya等,1992年;Molofsky & Fisher 1993)。
病虫害
在刚发芽的幼苗中,衰减是导致死亡的常见原因。潮气是一种由土壤真菌引起的疾病。Augspurger和Kelly(1984)通过野外和阴室实验对其进行了研究。在巴罗科罗拉多岛,新发芽的秀丽隐杆扁桃种子遭受了密度和距离依赖的死亡。将种子放置在密度为4倍的森林中,更容易发生水分流失,靠近成年树木的种子也是如此。以Barro Colorado岛18种风散树种为材料,在模拟小间隙(PAR为300 mol m ' s)和下层(PAR为17.5 mol m ' s)两种密度的遮荫室内播种。不同物种对阻尼的敏感性差异很大。研究发现,光线比种子密度更能决定种子衰减的可能性,在荫凉处死亡率最高。人群中可能存在许多其他疾病热带树木,但在这一领域开展的工作相对较少。据报道,象牙海岸的Turraeanthus africana (Alexandre 1977)和法属圭亚那的Dicorynia guianensis (Queneherve et al. 1996)的幼苗根部有潜在致病性线虫。
干旱
幼苗无法长出很深的根,这使它们特别容易受到干旱的影响。干旱期的高幼苗死亡率已被广泛报道(Turner 1990a)。在Barro Colorado岛的旱季,Virola nobilis幼苗在下层的死亡率最高(Fisher et al. 1991)。灌溉的幼苗在干旱导致较高的成活率。尽管有更大的蒸腾负荷,但缝隙中的幼苗在旱季中存活得最好,因为它们可以在缝隙中更高的光照下生长得更快,并发育出更广泛的根系,使它们能够在旱季中更好地获得水分。
阴影
大多数物种的幼苗在森林下层很深的阴影中生存和生长都减少了。这是本章后面一节的主题。就目前而言,只要说森林下层的低光照可能会造成如此低的光合速率,以至于幼苗几乎没有产生积极的影响就足够了碳排放增加.几天阴天可能就足以耗尽资源并杀死幼苗。
竞争
大多数幼苗生长在森林的下层,尽管周围有许多其他个体,但它们必须在那里生根发芽,其中大多数个体都比它们大得多。这是一个高度竞争的环境,优势竞争者可能会剥夺弱势个体的资源,降低他们的增长速度,甚至可能完全饿死他们。竞争导致的资源短缺,以及其他方式,将增加幼苗对疾病的易感性。靠近林下棕榈树对种植在森林林下的两种印加树幼苗的存活率产生了负面影响(Denslow et al. 1991)。Coomes & Grubb (1998a)发现,在委内瑞拉非常贫瘠的caatinga森林中,挖沟对幼苗的高度生长和叶片产量有积极影响。在下层和缝隙处,挖沟的响应幅度相似(图5.11)。
大多数树种的种子和幼苗死亡率都很高,再加上繁殖力和传播能力的不足,可能意味着许多树种在森林中是有限的。
继续阅读:耐荫性性状
这篇文章有用吗?