种子大小
比较生态学家对种子的大小很感兴趣,因为不同物种的种子大小差异很大。在热带物种中,种子的干质量至少超过六个数量级雨林的树.Melastomataceae和Rubiaceae包括热带树种,种子干质量只有20克(Metcalfe & Grubb 1995;Grubb & Metcalfe 1996)。在另一个极端,一些树木的种子,特别是豆类,接近100克干质量。在任何热带森林地点,大多数研究表明,树木种子质量的范围至少有五个数量级(Hammond & Brown 1995;梅特卡夫&格鲁布1995;Grubb & Coomes 1997;Lord et al. 1997)。当然,质量是一种与体积有关的属性,因此会随着所涉及的线性维度的立方而增加,这将迅速夸大物种之间的大小差异,但与动物群体相比,后代大小的百万倍范围仍然是巨大的。
种子大小可能受其他性状的异速生长控制。有证据表明,与其他大小变量相关。小植物产生大种子和小果实含有大种子的困难,可能总是导致植物大小和种子大小在某种程度上呈正相关(图5.1)。为热带树木在美国,一些研究表明,在特定的森林中,种子大小随成虫身高的增加而增加(Hilty 1980;福斯特&詹森1985;梅特卡夫&格鲁布1995;Hammond & Brown 1995;凯利1995年;Grubb & Coomes 1997)。散布机制也可能影响种子大小(图5.1),这可能是因为果实大小的分散剂选择。在三个新热带地区,哺乳动物传播的种子平均比鸟类传播的种子重12-14倍(Hammond & Brown 1995)。这可能是由于哺乳动物在森林中选择了水果大小范围内较大的一端,这似乎与进化有某种关系。然而,Kelly(1995)发现,在秘鲁的一个地点,这种模式经常在不同属中重复,这表明在哺乳动物分散的物种中,大种子大小的反复进化。
热带雨林的一个特点是存在种子很大的物种
-
 and successional status (filled, non-pioneers; open, pioneers). Data from Foster & Janson (1985).")
- 图5.1秘鲁马努树种种子质量与树高的关系。按传播方式分类的物种(圆形,哺乳动物分散;三角形,鸟类分散)和连续状态(填充,非先锋;开放、先锋)。数据来自Foster & Janson(1985)。
 and successional status (filled, non-pioneers; open, pioneers). Data from Foster & Janson (1985).")
(Lord et al. 1997)。Foster(1986)在综述大种子的适应价值时,提出了几个优点。然而,批判性分析并不支持接受所有这些观点。福斯特提出了以下大种子可能带来的好处。
延长种子寿命
简单地说,有人可能会说,越大的种子储备就越多,因此预计存活的时间也就越长。Foster(1986)在Ng的马来西亚树种大数据集中指出了发芽时间和种子大小之间的显著正相关关系(Ng 1980)。然而,Hopkins & Graham(1987)发现,在昆士兰州,埋藏长达2年的种子大多是小种子(图5.2)。Kanzaki et al.(1997)也发现热带植物种子大小与存活时间呈显著负相关森林土马来西亚的树种。对一些热带种子的呼吸速率测量揭示了这一悖论。Garwood & Lighton(1990)发现种子含水量对耗氧率的影响大于种子大小。休眠的种子含水量低,因此呼吸速率低。大的种子往往含水量高,因此呼吸迅速。对于含水量相似的种子,大种子具有较高的绝对呼吸速率,但单位干质量的呼吸速率通常较低。从理论上讲,能够干燥进入休眠状态的大种子应该能够存活很长时间,但这似乎是一种相对罕见的策略。大的休眠种子可能特别容易受到森林中脊椎动物种子捕食者的影响。
二次化合物的空间更大
大的种子可能储存更多的化学防御物质。这些可能被用来保护种子免受病原体或小型无脊椎动物的攻击。然而,对于大型食籽动物来说,化学防御的总浓度更有可能对食物选择产生影响。然而,非常大的种子可能含有比保险箱更多的物质
-
, hollow symbols, secondary forest species (mostly small seeds). Data from Hopkins & Graham (1987).")
- 图5.2来自澳大利亚昆士兰的物种种子在土壤中埋埋时间的变化。实心符号表示原生林种(多为大种子),空心符号表示次生林种(多为小种子)。数据来自Hopkins & Graham(1987)。
, hollow symbols, secondary forest species (mostly small seeds). Data from Hopkins & Graham (1987).")
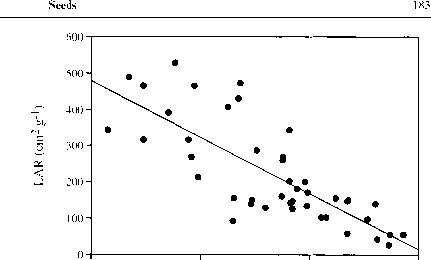
图5.3 43种热带树种小幼苗的LAR与种子干质量的函数关系。数据来自ter Steege (1994a), Osunkoya et al.(1994)和Kitajima(1994)。
种子干质量(g)
图5.3 43种热带树种小幼苗的LAR与种子干质量的函数关系。数据来自ter Steege (1994a), Osunkoya et al.(1994)和Kitajima(1994)。
每只动物每次只吃一部分毒素,从而提高了胚胎存活的几率。
增加光合作用组织与非光合作用组织的比率种子储量大可以使新发芽的幼苗更快地发育成更大的光合作用区域,从而有更大的机会在阴暗处生存。但事实似乎并非如此。Kitajima(1992)发现热带树种新发芽幼苗的种子质量与相对光合能力之间存在很强的负相关关系。小种子往往长出面积较大、子叶较薄的幼苗(图5.3),这使得小种子的相对生长率大大提高。微小的热带树苗能够在很深的阴影中持续生长(Ellison等人,1993年;Metcalfe & Grubb 1997)。
生长:在更高的光照下或更深的土壤中生长
更大的种子产生更大的幼苗(Janos 1980;Howe & Richter 1982),并且在更大的幼苗尺寸上可能具有优势。上面的光可用性可能有很强的梯度森林地面以及地下的可水量。大的种子可以长出更高的幼苗,得到更多的光,或者根深的幼苗得到更多的水。来自大种子的幼苗可能在更深的土壤或凋落物下成功发芽,并且可能更有力地与邻居竞争,更成功地生长菌根(Janos 1980)。在不利的时期,如被落叶遮蔽时,或在多云的时期,更多的储备将使更大的生存机会。Boot(1996)发现圭亚那六种树种在黑暗中发芽的幼苗在一年以上的存活率与种子大小呈正相关。在澳大利亚昆士兰州,大种子物种(如Castanospermum australe)的萌芽可以在掩埋中存活两年(Hopkins & Graham 1987)。
对损伤或组织损失的耐受力
由病原体、食草动物和掉落的碎片造成的损害可能会影响森林中种子和幼苗的生存。较大的种子储备有助于应对这些变化。就幼苗存活和高度生长而言,来自新几内亚的两个超大种子物种(新鲜质量超过100克)比其他两个种子大小小一个量级的物种更能容忍从种子中去除50%以上的非胚组织(Mack 1998)。大种子的碎片也可能发育成小株,如古斯塔维亚(Gustavia superba) (Harms et al. 1997)。Harms & Dalling(1997)测量了13种种子新鲜质量在0.2-107.6 g范围内的幼苗从斩首中恢复的能力。只有种子质量大于或等于5 g的5个品种在脱头后存活并再生。他们都有下萌发。
证据表明,大种子是有利的,因为它能产生较大的幼苗,其储量足以满足物理和化学防御、资源短缺时期和修复损害的要求(Kitajima 1996年)。热带雨林中有大量种子很大的树种,这可能反映了森林下层的阴影很深,竞争激烈,病虫害的威胁无处不在。在热带雨林中,潜在的大体型种子传播者的存在也可能有利于大种子。
然而,许多物种的种子仍然很小,这可能是因为种子大小和种子数量之间的基本进化权衡。如果一棵树有一定数量的资源可以用于繁殖,那么为了产生更多的种子,它就必须减少用于每颗种子的资源。更多的种子将意味着更多的尝试在下一代中产生可存活的后代。但一般来说,体型减小意味着死亡风险增加。因此,自然选择在不断增加的尝试次数和不断减少的生存机会之间进行平衡,并产生适合该物种的种子大小。从本质上讲,小种子风险更大,但每单位投资可以产出更多。
种子大小与种子数量之间的权衡可能不像前面一段看起来那么直接,至少就肉质水果而言。种子小的果实果肉干质量与种子干质量之比较高(Grubb 1998b)。这可能是因为小水果比大水果需要按比例提供更大的奖励来吸引分散者。
在某些情况下,种子的大小与生存关系不大,或者小种子具有特殊的优势。有人认为,在资源相对丰富的林隙环境中,种子大小对幼苗存活的影响可能较小,而小种子产生的幼苗由于相对生长速度较高,可能会迅速赶上大种子产生的幼苗。这种观点认为强烈需要光照的物种的种子比那些在这种观点在生态学文献中根深蒂固(Swaine & Whitmore 1988),但最近受到了挑战(Kelly & Purvis 1993;Grubb & Metcalfe 1996)。比较被认为需要间隙再生的物种和那些可以作为幼体在森林下层存活的物种之间的种子大小,通常表明热带雨林中耐荫性的平均种子质量(或其他大小指标)更大(Foster & Janson 1985;Hammond & Brown 1995),但这总是涉及到两个物种组的种子大小非常大的范围。然而,校正可能的系统发育影响的统计分析显示,在耐荫性中,更大的种子大小的证据要少得多(Kelly & Purvis 1993;Grubb & Metcalfe 1996)。Grubb和Metcalfe(1996)认为,这些分析暗示了种子大小有很强的系统发育影响,表明这种特征表现出进化惯性。在一个特定的分支中,种子的大小在进化过程中变化缓慢,对于一个物种是否能成功地成为一棵耐阴或需要间隙的树可能没有特别的影响。大种子的光需求植物包括Aleurites moluccana(7.8克干质量)(Grubb & Metcalfe 1996)和Ricinodendron heudelotii(1.4克干质量)(Kyereh et al. 1999)。各种作者(Metcalfe & Grubb 1995; Grubb 1996; Grubb & Metcalfe 1996; Hammond & Brown 1995) have highlighted the presence of very small-seeded, strongly shade-tolerant species among rain-forest tree floras. Hammond & Brown (1995) hypothesised that small adult stature, low resource availability in the forest understorey and use of small-bodied dispersers compound to favour small seed size. These are likely to be selection pressures acting in favour of a trend of decreasing seed size with distance below the top of the canopy, but they are probably still insufficient to explain the very tiny seeds of some species, notably among the Rubiaceae and Melastomataceae. These minuscule seeds are possibly specialist exploiters of certain regeneration sites in the forest. Given the small size of the seeds and their subsequent seedlings, regeneration could not be successful if litter were present, so litter-free microsites on the forest floor are required. Sites on steep banks and slopes are most suitable, where large seeds would fall down, but the minute seeds are caught by tiny irregularities in the soil surface (Grubb & Metcalfe 1996; Metcalfe et al. 1998). Grubb & Metcalfe (1996) point to the possibility that these species have evolved from light-demanding ancestors.
委内瑞拉caatinga的高大树木的种子比附近肥沃土壤上的类似树木的种子更小(Grubb & Coomes 1997)。这被解释为在土壤养分贫乏减少了可用于繁殖的资源的环境中,猫鼬物种保持了种子数量。同一组作者认为,在这样的低肥力地点,根系竞争更加激烈(Coomes & Grubb 1998a),人们可能会预测这将有利于较大的种子物种,但增加种子大小可能带来的好处增长太慢,不足以抵消产生较少种子的缺点。
继续阅读:种子发芽
这篇文章有用吗?
读者的问题
-
luukas3个月前
- 回复