The traits of shade tolerance
幼苗的反应shading are similar across a wide range of species. Increased allocation to shoots, particularly leaves, the production of leaves of larger area but thinner laminas, and reduced dark respiration and maximal photosynthetic rates, make up the nearly universal shade response. Why, then, cannot all species adjust physiologically to cope with very low light availabilities? The answer seems to be that all but a few can. A small proportion of species appears to be unable to reduce respiration and leaf production and turnover rates to cope with deep shade. Such species enter a negative carbon budget, because high respiration and the inefficient turnover of leaves are not balanced by photosynthetic gains, and soon expire, although often they succumb to disease before they have respired all their resources. If the definition of shade tolerance as the relative ability of populations to persist in the deep shade of the forest understorey is accepted, then survival at low intensities ofincident radiationbecomes the key feature of shade tolerance. An ability to withstand damage and disease at low light availabilities is a more important trait of shade tolerance than maximising photosynthetic gains. Kitajima (1994) found that leaf gas exchange rate had few significant correlations to seedling survival in the shade for 14 species from Barro Colorado Island, Panama. Shade-tolerant species (ones more persistent in shade) had greater LMA, higher root: shoot dry mass and lower LAR in the shade than less tolerant species. This is exactly the opposite pattern to the general response to increasing shade. In other words, more shade-tolerant species show less morphological and physiological plasticity to shading than more shade-intolerant species (see, for example, Strauss-Debenedetti & Bazzaz 1991; Ducrey 1992; Strauss-Debenedetti & Berlyn 1994; Veenendal et al. 1996; Veneklaas & Poorter 1998). An exception to this pattern was noted by King (1991b), who found that saplings 1-2.5 m tall of species strongly associated with gaps varied little in shoot dry mass allocation to leaves with light conditions whereas shade-tolerant species increased allocation to leaves with reduced light availability.
Kitajima解释的一般模式that these are features necessary to improve survival in the deep shade. Shade-tolerant species allocate more to defence and long-term security than shade-intolerant species, which show greater plasticity in form and physiology. The shade-intolerant species end up with a highly 'risky' seedling design: very thin leaves are susceptible to herbivores, the limited root system results in sensitivity to drought, and so on. Evidence for a seedling survival versus growth rate trade-offwas found among 14 pioneer species on Barro Colorado Island (Dalling et al. 1998a).
Shade-tolerant species, in a pattern similar to plants typical of other habitats poor in available resources, tolerate shade by harbouring their reserves, protect themselves from potential enemies and emphasise survival rather than maximal growth rates. The higher LMA of shade-tolerant species in the shade probably reflects greaterleaf longevity(a longer time over which the leaf can make profits for the plant). The steeper slope for the correlation between leaf maximum assimilation rate and foliar nitrogen or phosphorus concentrations observed in shade-intolerant species (Strauss-Debenedetti & Bazzaz 1991; Raaimakers et al. 1995) may reflect the allocation of resources to processes other than photosynthesis, such as defence in shade-tolerant species. Pioneers typically have greater instantaneous assimilation rates per unit of nutrient in the leaf (see Chapter 2).
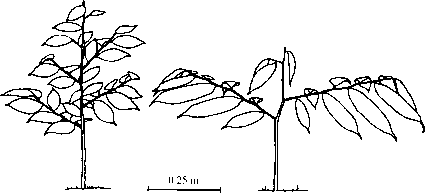
There may also be architectural differences between the two shade-tolerance categories. It has been argued that light-demanding species tend to be strongly orthotropic, with the leader shoot showing strong apical dominance, producing tall and thin seedling crowns. Shade-tolerants have a greater tendency to be plagiotropic, with spreading branches producing sprays of leaves and a wider crown. This dichotomy of form is exemplified by the seedlings of two dipterocarp species, Dryobalanops lanceolata and Shorea leprosula (Fig. 5.14). Kohyama (1987) named the species following these architectural strategies optimists and pessimists, respectively. This is because the orthotropic species behave in an optimistic manner, emphasising height growth into higher light conditions. The plagiotropic species are pessimistic, spreading sideways to capture more light and allow them to persist in the shade. However, Kohyama & Hotta (1990) found that tropical seedlings showed a large diversity in architecture and the pessimist-optimist dichotomy may be too simplistic to apply there.
A number of studies have reported that wood density (usually adult wood density obtained from references on timber properties) is a positive correlate of shade tolerance in juveniles (Augspurger 1984; Whitmore & Brown 1996). There are several possible reasons for this. Very fast-growing species rarely have dense wood, probably because such growth rates could not be achieved if more resources went into the wood. Dense wood may be of positive value to plants in the understorey. Wood density is correlated with resistance to decay by fungi, and probably also to wood-boring insects. Wood strength is correlated with density and the understorey is a place where wood strength may mean the difference between death and survival when a falling branch lands on a seedling.
Shorea leprosula Dryobalanops lanceolata
Figure 5.14 Seedling architecture of Shorea leprosula and Dryobalanops lanceolata. Dryobalanops shows preferential resource allocation to the growth of a few long branches. Individuals shown have the same age and total branch length. After Zipperlen & Press (1996).
Shorea leprosula Dryobalanops lanceolata
Figure 5.14 Seedling architecture of Shorea leprosula and Dryobalanops lanceolata. Dryobalanops shows preferential resource allocation to the growth of a few long branches. Individuals shown have the same age and total branch length. After Zipperlen & Press (1996).
The denser, stronger wood of the understorey species would allow them to have narrower stems for a given loading. However, the generally more spreading crowns of the juveniles of understorey species might mean that they would need proportionally thicker stems for a given sapling height than large-statured species. These two factors acting in opposite directions may help explain some apparent anomalies obtained in allometric analyses of sapling form for tropicalrain-forest treespecies. King (1994) studied the allometric relationships of six species in Panama and found that the two species of smallest adult stature had significantly thicker stems with wider, leafier crowns at 2.5 m height than the rest. Both Thomas (1996a) and Davies et al. (1998) reported that saplings of species with low asymptotic height tended to have narrower stems as saplings than those of species of greater maximal size. The latter study concerned 11 species of the mostly strongly light-demanding genus Macaranga, where the small-statured shade-tolerants were unbranched treelets with narrow, rather than wide, crowns. Wood density differences may have allowed the small-sized species studied by Thomas not to differ significantly in trunk (H-D) allometry at the sapling stage.
The orthotropic unbranched, or sparsely branched sapling architecture seems to have advantages when it comes to rapid height growth. Coomes & Grubb (1998b) found that the juveniles of tall-tree species with this architec ture had the fastest relative height growth rates in tree-fall gaps of species studied in the caatinga forest of Venezuela. The erect unbranched form allows greater height increment per unit increment in dry mass (King 1994).
Continue reading here:BOX 61 The nomenclature of the pioneerclimax dichotomy
Was this article helpful?