树木形成力学和水力学树形象
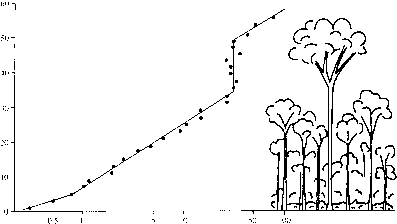
个人树大范围的大小是在热带雨林中找到(请参见图2.1中的概要图)。每个树种也有到期特征尺寸和物种通常被称为各种身材类,如下层木树,树冠树木和紧急,但是,如图2.2所示,没有离散集群大小类的物种。最大直径Pasoh物种50公顷的森林,马来西亚半岛(图2.2),大约是与模态截断对数正态分布分布类最大胸径范围在10至20厘米。崩溃的物种为高度类(表2.1)显示大约一半的树种有最大20米的高度,或更少。最高的树在Pasoh可能达到约60米高,表明下层木专家的社区在森林里一样丰富的树种树冠,单位深度基础上和绝对是丰富的。在森林里一个类似的模式在热带雨林,哥斯达黎加(鹿角1980),和巴罗科罗拉多岛,巴拿马(哈贝尔&培养1992)。
树种多样性的垂直分布在森林里可能反映了在不同高度高于相对照明森林地面。在Pasoh,相对光照随高度增加对数(图2.3)。下层木物种在底部20米的森林很少获得超过5%的辐射到达顶部的树冠。然而,与高度的变化率相对照明是最大的森林附近的地板上。换句话说,对于一个给定的高度相对增加,而不是绝对的,增加照明更大的森林地板。也许这陡坡促进分区物种间的光资源,因此单位深度比更高的专家更多物种在森林里。
已经被越来越多的趋势在热带森林文献使用术语灌木和treelet归类物种的最大高度,与“灌木”代表着最小的树和“treelet”下一个大小类,两者之间任意设定限制。更传统,灌木也杰出小木本植物与多个茎,或分支十分贴近地面。声称,“真正的灌木是罕见的热带雨林。Givnish(1984)指出,只有6 95种non-climbing伍迪的不到10米高成熟度原生加纳是真正的灌木而温带森林下层木经常有许多灌木的物种。multi-stemmed皇冠建设可能是更有效的方式支持低,广泛的树冠比单个干细胞与深远的分支。还没有强烈支持假设为什么灌木是罕见的雨林社区的成员。
treelet是生命形式可能比其他任何平民在热带雨林植被类型:single-stemmed、无支链的或稀疏分支树到期矮小的身材。达奇(1973)提出的两个主要treelets的进化起源。他们在伍迪组织起来通过早熟的开花,或通过伍迪茎的发展否则草本进化路线。后者是在夏威夷,在那里种Cyanea和森林treelets Plantago发生。

0 10 20 30 40 50 60
图2.1概要图Ulu Dapoi、森林的Tinjar,婆罗洲。一块60 m x 8 m (200 x 25英尺)。注:艾什顿教授提供的资料。
0 10 20 30 40 50 60
图2.1概要图Ulu Dapoi、森林的Tinjar,婆罗洲。一块60 m x 8 m (200 x 25英尺)。注:艾什顿教授提供的资料。
身材 |
定义 |
不。的物种 |
所有物种的比例(%) |
树木每公顷 |
茎的比例(%) |
“灌木” |
2米高 |
54 |
6.6 |
303年 |
4.6 |
“treelet” |
2 - 10米高 |
雷竞技csgo |
13.5 |
1513年 |
22.9 |
下层木 |
10 - 20米 |
283年 |
34.5 |
2114年 |
32.1 |
树冠 |
20 - 30米 |
317年 |
38.7 |
1901年 |
28.9 |
紧急 |
> 30米高 |
55 |
6.7 |
753年 |
11.4 |
数据从Kochummen et al . (1990)。60-1 -
数据从Kochummen et al . (1990)。60-1 -
0 -H-v-LrJ——T”^先生^ -我我我我,v
观察到的最大胸径(厘米)
图2.2分布最大树干直径的树种有超过300个人在Pasoh 50公顷的情节,马来西亚。后Kohyama (1996)。
我将用更确切的灌木和treelet形态意义的文本,并参考侏儒树木当讨论最小的树状分类单元无论架构。
在巴罗科罗拉多岛,最常见的物种在50公顷的阴谋treelet Hybanthusprunifolius,与近40 000人238 000枚举茎。或许有人会说这是侏儒树更容易是常见的,因为它们更小,因此更多的人
挤在一个区域。这忽视了一个事实:有小少年的大树,这使得它理论上最大的树种有最大的人口密度。然而,small-statured物种必须有最大的空间密度高繁殖成熟个体。汉堡(1980)指出,侏儒在哥斯达黎加雨林的树木往往是来自Piper等大型属和Psychotria,特别是更多的物种属的植物比更大的树。泛热带范围内,这两个庞大的灌木的属被他人管理和Rinorea等加入,虽然需要一个更详细的分析证实,雨林侏儒树通常有一个更大的物种属比更大的生命形式。
木
木头是派生的次生加厚的根和茎轴。次生加厚涉及细胞的穹状的部门。这发生在一个外围形成层在典型的树,但在分散网站通过轴在许多树状的单子叶植物。功能、木材包含三个主要元素:木质部导管(相似和/或船只)进行木质部汁液,木质部纤维提供大部分的机械支持(裸子植物木材的传导和支持的作用不太明显划分为相似
的意思是相对照度(%)
图2.3垂直剖面的平均相对照度(表示为一个百分比的光树冠上方)在雨林网站Pasoh、马来西亚。后,理查兹(1996)。
的意思是相对照度(%)
图2.3的垂直廓线意味着相对照度(表示为一个百分比的光接收树冠上方)在雨林站点在Pasoh,马来西亚。后,理查兹(1996)。
单独负责)和薄壁组织细胞,可能是重要的横向传导的水和存储,并可能帮助渠道从空化中恢复过来。前两个细胞类型非常明显拉长的方向长轴的组织,木材的纹理,渠道提供管道工程水沿着茎或根的运动。
木材是一种优良的结构材料(Jeronimidis 1980),因为它结合了强度和刚度和高阻的传播裂缝相对较轻的复合显示的温度依赖性的力学性能相对较少。然而,它是一个高度各向异性材料。其抗压强度一般是一半的抗拉强度,强度横纹是10到100倍不到一起,骨折(韧性)和工作只是高横纹。
长列细胞,尤其是纤维相似,提供木材顺纹的力量,和木材的抗裂纹扩展它。射线举行轴向纤维一起抵制tor-sional和剪切应力,确定树的横向强度。木材细胞纤维素微纤维沿长轴成螺旋形地运行S2细胞的壁(图2.4)。微纤维的方向大约25°从细胞的长轴垂直的优化他们的贡献,木材的抗拉和抗压强度。木质素中扮演一个重要的角色在交联并巩固细胞壁的纤维素成分从而提高木材的刚度和强度。S2微纤维也耐开裂的重要贡献者木材横纹。避免裂纹扩展是很重要的,因为所带来的结构性失败通常是裂纹形成局部地点的高应力强度而不是力量超过材料的强度。卢卡斯et al。(1997)提出,在裂纹尖端的应力集中传播在木纹会导致内心崩溃的S2-microfibril层纤维细胞。微纤维拉墙进入腔沿着几乎完整的细胞的长度。这个崩溃的纤维层代表了相当大的能量的使用,和塑性变形可能高达90%的负责工作期间断裂的木材。
木材的力学性能是强烈与其密度呈正相关。木材的实际固体组件的密度变化小树种之一,被约1.5厘米~ 3 g风干物质(威廉姆森1984;Detienne &小调1996)。是空间内和细胞之间的存在导致木材密度的变化。低密度的木材,如巴尔沙(巴尔沙树pyramidale),与大型薄壁木材细胞腔和丰富的细胞之间的空间。最密集的心材木材方法空间密度最大。强度属性是直接关系到固体材料的数量,但木材的细胞性质导致裂纹阻力(卢卡斯et al . 1997年)。非常密集的木材很少,如果有的话,纤维内腔。因此塑性变形的数量可能减少开裂和断裂的工作期间下降。因此非常密集的木材,如愈疮树(愈创spp。),是很难和强大,但容易脆弱。 Very low-density timbers have lower toughness than expected because their thin cell walls fracture before they can collapse.
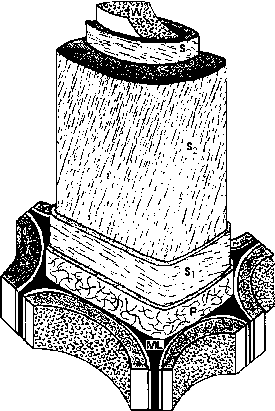
图2.4超微结构的示意图表示木材纤维墙的组成部分。毫升,中层,P,初生壁,年代,次生壁,呈现为三层,S ^ S2和S3。S2墙上包含螺旋绕组纤维素微纤维。W代表有疣的内腔的表面。在一些物种中表面是光滑的。
图2.4超微结构的示意图表示木材纤维墙的组成部分。毫升,中层,P,初生壁,年代,次生壁,呈现为三层,S ^ S2和S3。S2墙上包含螺旋绕组纤维素微纤维。W代表有疣的内腔的表面。在一些物种中表面是光滑的。
热带木材
木材的显著特征热带树木宽的存在(大内部直径)容器元素,通常使用简单的穿孔板(Carlquist 1988)。大血管的主要优点是增加水传导率,因为流体流经管道的利率与四次方的直径成正比。然而,并不是所有的船只在热带木材宽。通常有许多直径较小的船只。这些可能提供安全通过冗余的导电系统树,和径向传导可能是重要的水(批et al . 1994年)。非常广泛的血管,超过100 | im直径,是罕见的在树上,常见的只有在液流率要求高的藤本植物,茎的横截面积有限。热带树木拥有更广泛的血管将允许更大的液流率通常通过树干和树枝,但当交付率的水的叶子不会增加电导率提高的阀杆和更高的流动阻力的主要分支,因为外面的树枝。宽的船只往往被认为是更容易比窄的空化。批&大口水壶(1996)发现了一个弱负相关之间施加压力要求导致渗透系数降低50%和平均血管直径大树种的选择。然而,更大的对空化机制在更广泛的木质部导管只有清晰的freezing-induced栓塞。 Large air bubbles take longer to dissolve than small ones, increasing the likelihood of serious cavitation after the freezing of xylem sap (Tyree et al. 1994). Freezing temperatures are not experienced in the lowland tropics, but they are in the temperate zone, where trees tend to have vessels of smaller diameter (van der Graaff & Baas 1974). This is probably why temperate timbers of the same density as tropical ones consistently have 20% lower cross-grain work of fracture (Lucas et al. 1997). The greater vessel number in the temperate woods reduces their mechanical performance. It is questionable, however, whether there has been selection for a certain vessel size in tropical trees to improve the mechanical properties of the wood. Roots tend to have wider vessels than stems (Ewers et al. 1997), probably because roots are less liable to cavitation and are better able to recover from it.
热带森林的木材密度的广泛存在,经常包括物种都很低(小于0.3 gcm ~ 3)和高(超过0.7 gcm ~ 3)密度(威廉姆森1984)。木材密度通常报道说重量每卷以12%的水分(雷耶斯et al . 1992;Detienne &小调1996),尽管生态用途真实密度、烘干的绿色质量/体积,更有用。雷耶斯et al。(1992)提供了一个回归方程进行密度估计的两种类型之间的转换。热带森林的多品种样本显示正常的木材密度分布(图2.5)与手段在0.560.62 g厘米(雷耶斯et al . 1992;Detienne &小调1996)。这些研究的结果提供支持惠特莫尔&席尔瓦(1990),南美,尤其是亚马逊,木材密度大,或者至少显示更大的密度有很重的传播伍兹现在,比其他热带地区。
木材密度与分类有亲和力。很轻的木棉科是著名的木材,特别是在Cavanillesia等属、木棉和巴尔沙树。等低密度森林中发现属Cecropia, Musanga,榕树,苹婆属。很重的森林往往从caesalpinoid豆类的亚科;Swartzia就是一个很好的例子。乌木(Diospyros柿科)也非常沉重的木材而闻名。
有证据表明,在热带森林的木材密度范围缩小与海拔和扩大与增加干旱的严重程度(威廉姆森1984)。树种发现更多接触网站在哥斯达黎加大部分山区森林木材密度高于更庇护的地方的物种特征(劳顿1984)。推测物种低密度、弱木将处于强烈的劣势更大的风荷载下的山的山脊。种drought-deciduous森林的木材解剖Chamela比较与洛Tuxtlas湿低地森林,还在墨西哥(Barajas-Morales 1985)。的常绿林树木有柔软、lighter-coloured和密度较低,木材用更少的水晶树脂夹杂物比落叶林的物种。平均血管直径较低从Chamela在树上,是典型的更多的季节性干旱森林(批et al . 1994年)。比较30种,生长在雨林和草原网站在科特迪瓦,洞外& van Veenendaal(1976)发现的大多数物种有更广泛的干燥草原地区的船只。网站之间的整体平均血管直径的差异没有统计学意义。
并不是所有的树木心材形式:比如手掌,以及快速增长的soft-wooded双子叶植物物种例如Dyera costulata,没有。这些边材树可以通过机构积极捍卫自己的木活细胞的整个躯干和很少,如果有的话,发现空心因为任何害虫和疾病成为建立可能会毁了整棵树。死者心材构成的核心大多数树木的树干只能被动行动免受腐蚀的成分和被覆盖的上覆住边材。
密度也可以衡量的程度伍迪器官免受攻击害虫和病原体。的力学性能
-
.")
- 图2.5频率分布的热带森林物种类木材密度三个热带地区。之后,雷耶斯et al。(1992)。
.")
木材在规模较小可能有助于防止腐烂。动物会发现非常密集,硬木难以分割摄入之前,和真菌菌丝将无法渗透和分解伍迪组织。然而,heartrot发生和空心树热带森林中并不少见。
木材密度经常被使用的森林和生态学家作为衡量一个物种的最大增长率和相对树荫下公差。快速发展,shade-intolerant物种木材密度较低,而木材生长缓慢,那些在物种是沉重的。令人惊奇的是竟没有可靠的定量证据来证明这个论断。木材密度显示明显,虽然弱负相关,平均直径增长速度在122个物种从法属圭亚那(Favrichon 1994)。在婆罗洲雨林,树杆比重(包括树皮)0.2 - -0.49表现出显著的速度相对茎直径增长比茎的比重超过0.5(铃木1999)。幼苗坚持深沉与成人木材密度表现出显著的正相关关系在18风树从Barro科罗拉多岛,巴拿马(Augspurger 1984)。木材密度是最好的(尽管仍然疲软)光的预测需求指数从昆士兰树种对再生,澳大利亚(Osunkoya 1996),但该指数的排名生态学家的印象,而不是一个可量化的衡量耐荫提出相关先验。
大部分的木材,纤维细胞相似,容器元素是死了,但射线薄壁组织和其他边材组织生活,因此呼吸。伍迪组织的呼吸率两个树种在热带雨林,哥斯达黎加,测量(Ryan et al . 1994年)。快速增长的Simarouba amara (1.24 | jmolm ~ 2 s“我)被发现呼吸速度大大快于缓慢Minquartia guianensis (0.83 | imol m ~ 2 s“我)。津贴为木材体积增长率时发现这两个物种有着差不多的维护呼吸率。这些利率是温带针叶林的两倍。热带树木的高维护呼吸可能是由于高温有经验。快速呼吸的主要原因,尽管增长条件优越,热带森林的木材生产率并不比在温带,甚至更大的北方森林(约1982)。
树木的机械设计
的一个关键限制树的形式是其必要的能力来支持自己的体重和范围外部力量产生的,尤其是那些风,很有可能在它的生命周期。Mattheck & Kubler(1995)提供了很好的概述树设计如何帮助解决这些机械问题。失败通常涉及局部区域的过度压力。树木可以概括为增长的机械设计和分配的材料保持一致树内的压力和避免关键强调在特定的地点。这是通过多种机制,包括:
1。被动弯曲灵活的部分。不反对外部力量最大限度地减少装运,但大型重型茎会扣在自己的体重如果他们非常灵活。因此,弯曲的策略是局限于小乔木,大树的终极分支,特别是竹子(Mosbrugger 1990)。Rheophytic树木和灌木,生存时间的洪水水流湍急的河流中使用他们的灵活性,和顽强的根系统,才能生存。树冠需要一些灵活性,减少对气流的流动阻力在有风的日子。沉重的风载荷树下可能产生的弯曲木(Ennos 1997),具有更明显的螺旋纹。它的密度比正常的木头,短,thicker-walled细胞。纤维素微纤维也伤口比平时更大的角度。从理论上讲,这应该会导致木材低刚度相同的断裂应变。 As yet, only the former has been demonstrated experimentally (Ennos 1997). Branches may have denser wood than tree trunks so that they can be relatively thinner and thus bend and 're-configure' more easily. Putz et al. (1983) noted that is was low-density-timber trees that tended to lose limbs in暴风在巴拿马。
2。木材的设计作为一个蜂窝复合材料。木材的各种组件排列满足不同机械的挑战。而纤维的轴向排列,一样提供阀杆的强度,射线纤维抵制临界剪切和扭转应力。单子叶植物的木材,其扩散厚壁材料,缺乏有效的十字支撑和不擅长抵抗力量的方向平行于主轴。这或许就是为什么树状的单子叶植物往往无支链的皇冠有限发展(Mosbrugger 1990)。分支插入和传播冠不可避免地产生侧压力。手掌克服这个限制在一定程度上通过巨大的树叶,相当于整个分支。这些支持的高纤维叶基地扣主干紧密。
3所示。树的外形和木材的内部质量优化,以满足外部力量。树干通常大约圆形截面。成角的茎是不利的,因为他们会导致高应力强度的角度。拱是使用增长的另一个例子来克服机械挑战(见下文)。显著的积极的木材密度梯度髓树皮被发现
16的20种树测试在哥斯达黎加(Wiemann &威廉姆森1989)。手掌也往往有密集的木的树干底部的外围(富1987)(图2.6)。Wiemann &威廉姆森(1989)发现,这些在低密度梯度最明显木材如木棉pentandra(图2.7)。这种树木的研究表明,木材密度梯度的变化所带来的木材生产的密度随着树的年龄,而不是局部的密度增长木材(达&威廉姆森1992;德卡斯特罗et al . 1993年)。树苗木材的快速增长Laetia procera有一半的成年树的密度,但树苗和成人森林林分Dipteryx panamensis是相同的密度(王1996)。有可能是高度发展的激烈竞争

图2.6密度分布示意图的干细胞成熟的棕榈树(椰子)。后Killmann (1983)。
半径(cm)
图2.6密度分布示意图的干细胞成熟的棕榈树(椰子)。后Killmann (1983)。
-
 of Ceiba pentandra. After Wiemann & Williamson (1989).")
- 图2.7比重作为距离的函数两棵树(不同的符号)的髓木棉pentandra。后Wiemann &威廉姆森(1989)。
 of Ceiba pentandra. After Wiemann & Williamson (1989).")
青少年light-wooded热带树木的好处很软的“低成本高的风险”使用木茎和分支。树干生产中的主要强调通过弯曲和发生在茎的最外层部分横截面。主要支持中央部分材料上面的重量。去除中央部分,即做一个树洞,改变树干的力学性能相对较少,只要外部实体墙的厚度的比值(t)空心的直径(D)仍然是相当大的。茎的削弱的中心柱由质量弥补失去的去除(Mosbrugger 1990)。中空茎机械意义和在一些快速增长的物种包括成员属Cecropia Musanga,其中t / D通常落在1/3到1/5不等。然而,工程与中空茎分支插入困难。空心分支会议空心杆往往导致高压力在插入点。Cecropia通常短,僵硬的分支机构将这些压力低,并且可能依赖于髓膨减少屈曲的风险更小的分支。大,老树通常有中空的树干。 They appear to be mechanically effective until t/D approaches a critical value of 0.3 (Mattheck & Kubler 1995).
4所示。生产反应木高应力强度的点。反应木裸子植物和双子叶树之间是不同的。松柏产生压缩木材,富含木质素和微纤丝角高S2墙壁的木材细胞。针叶树木材的反应有高强度产生压缩和高压缩的网站底部等主要的四肢。双子叶植物产生张力木,小木素和低microfibrillar角。它是生长在网站等高压压力上的大树枝。
5。内部木筋预应力平衡临界载荷。有证据表明,预应力树干,树干下的表面张力和压缩下其核心。这提高了树的性能对其最有可能的一般模式失败:产生的压应力通过弯曲的树的方向弯曲(背风的一面,如果是风造成弯曲)。如果这个地区的木材开始下张力需要更大的力达到临界压缩比树干无筋预应力加载。予将意味着更高的紧张压力在树干的迎风面,但木材强度比抗压强度。
拱
低地热带森林的突出特点是支持根的相对较高的频率从大树的基础。在森林调查的树木在马来半岛,41%的个人胸径45厘米或更大的拱为1.3米或更高(Setten 1953)。37%的图被发现同样的树木大小和buttress-height团体在委内瑞拉(Rollet 1969)。十八岁的78种(23%)超过10厘米胸径在Kibale拱在乌干达(查普曼et al . 1998年),与大类倾向于有更高频率的支撑。拱在大小和形状变量之间的物种,在一些非常高。例如,Rollet(1969)报道Sloanea guianensis和Ceibapentandra让他们扩展到超过7米的树。一些热带树种几乎总是发现拱(88%的Intsia palembanica树(Setten 1953)调查),他们几乎从不有一些属(0.8%),和其他支撑网站和基因型有不同的看法。常用的支撑形式帮助森林植物物种鉴定手册显示强大的遗传控制支持设计。
工程参数经常提出解释为什么树需要支撑。他们的名字,来自建筑设备用来保持建筑站,意味着机械功能。然而,直到最近,物理学的原理应用于树木为了发展支撑的功能解释树(Mattheck 1993)。拱的基本前提是树的机制有效地传输到地面荷载而引起的应力由于风(Ennos 1993 a)。永久安克雷奇树需要传输的压力。如果不是那棵树将会下降。树深和厚直根立即下树干可以直接向地面传输力量。然而,许多树木的最大根辐射的树干底部的上层土壤,创建一个浅盘伍迪的根。这些根板很容易观察wind-thrown树热带雨林。伸卡球根,降板深入土壤,传输中产生的压力的地上部分树在地上。Mattheck提出,会产生很大的力量之间的连接的树干底部和侧根树时受到风荷载。这样的部队很容易折断或分裂与主干的根结。应对这些压力的浓度,少年树主干和主要横向根之间的发展扩展,最终成为拱。这些支架侧压力的树干和允许更有效的传输和降低应力集中和根树干之间的连接基地。
拱可以作为张力和压缩元素取决于风的方向(图2.8)。迎风一侧的树的树干弯曲倾向于把根从地上。拱传递这种紧张顺利的根,像绳索。根系被推到地面上生成的压缩力量背风一侧的树。这里的拱作为道具蔓延出压力。Ennos(1995)电子应变仪用来证明内应变的定性模式支撑树Mattheck曾预测,最高的菌株在发展中桥墩的顶部。
Mattheck最好的实验测试的假设拱进行了功能意义的小树在低地雨林丹浓谷,沙巴,马来西亚(骗子et al . 1997年)。两组的树木进行了研究。Mallotus wrayi代表一个典型unbuttressed物种。鉴定了总共pinnate-leaved,通常支持个变种被发现包括个人的几个种类的光之女神和综述ramboutan-ake。支撑树并不总是伸卡球根,他们有时tap-roots。树木受到严重的仿制品
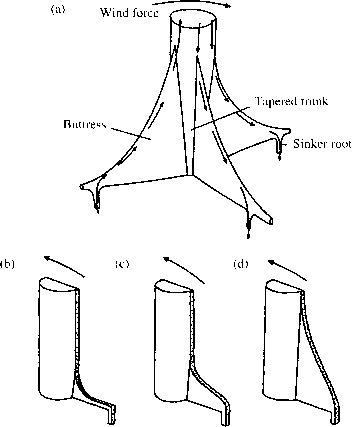
图2.8 Mattheck桥墩模型的功能和发展。如果一棵树被风(a)在弯曲力传播顺利拱的侧向伸卡球根。迎风向上下坠球抵抗力量和支撑放在紧张而背风下坠球抵制向下的力量和拱压缩。部分(b)、(c)和(d)显示连续Mattheck模拟的巩固发展阶段。当主干是拉应力集中的顶部结侧根和树干之间的点彩。增长的地区(b)的压力很大导致拱的形成(c, d)和一个伟大的浓度减少压力。后Ennos (1995)。
图2.8 Mattheck桥墩模型的功能和发展。如果一棵树被风(a)在弯曲力传播顺利拱的侧向伸卡球根。迎风向上下坠球抵抗力量和支撑放在紧张而背风下坠球抵制向下的力量和拱压缩。部分(b)、(c)和(d)显示连续Mattheck模拟的巩固发展阶段。当主干是拉应力集中的顶部结侧根和树干之间的点彩。增长的地区(b)的压力很大导致拱的形成(c, d)和一个伟大的浓度减少压力。后Ennos (1995)。
风载荷的绞车。在支撑树没有下坠球,失败是背风侧根的破损和退出迎风的(图2.9)。随着下坠球,迎风侧根分层。横向根高矩形截面,高八倍宽。这有助于根部抵抗弯曲向上或向下。侧根相对无效的锚定在non-buttressed树。侧根退出,或弯曲压缩,容易。两倍的支持个人稳定的unbuttressed树大小相同。研究证实了Mattheck的预测,拱可以有效张力和压缩两大元素,阻止根被拉出地面或推。存在的下坠球拱在张力大大提高了它们的有效性,但即使没有下坠球他们工作在压缩。 Possibly, as the trees grow larger more sinker roots develop. Reaction wood is not commonly found in the buttresses of tropical dicotyledonous trees (Fisher 1982). The increased tensile strength of reaction wood would possibly carry a disadvantage of reduced compressive strength.
考夫曼(1988)认为,支持发展危机的形成很大程度上是由于在树上的生活由于皇冠不对称等因素或突然暴露在大风后差距形成或增长主要的树冠之上。树上的支持可能会持续,尽管减少导致其最初形成的因素。支持这个理论,查普曼et al。(1998)发现火情物种在基巴莱,乌干达,显示支撑明显低于树冠和紧急物种,即使修正为个体大小。
皇冠不对称可能在热带雨林的树木很常见。127棵树的平均程度的皇冠不对称(大于20厘米胸径)观察到巴罗科罗拉多岛,四分之三的皇冠的一个典型的树是偏胖的年轻&哈贝尔(1991)。树木往往冠发展为空白或远离大的邻国。紧急通常是比个人小身材更匀称。在6.7年越不对称个人有更大的几率下降比对称冠,他们倾向于下跌对重侧(年轻& Perkocha 1994)。扶壁形成在树上是更明显的主干与沉重的皇冠。大树,桥墩通常比总树大小规模增长更快(年轻& Perkocha 1994)。这将减少的有效长度伯乐树有关,可能有助于降低躯干屈曲的风险。拱使树硬的基础和可能允许更高的高度和更广泛的冠实现相对安全。
拱的另一个可能的优点是降低要求
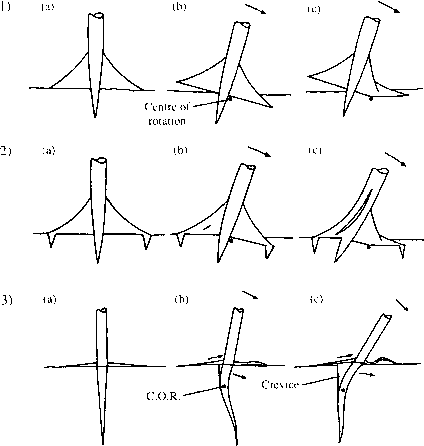
图2.9树干和根运动在安克雷奇失败的支撑和non-buttressed树。(1)支撑树没有伸卡球根(光之女神竹),(2)支撑树伸卡球根综述ramboutan-ake)。注意:伸卡球根可能存在或没有光之女神和综述的物种。(1)支撑树没有伸卡球根。树是由厚横向支撑固定在地上的树根和直根。(1 b)作为树拉,躯干旋转点只是背风的一面。最初,根树牢牢地固定在地上,背风侧抗弯曲被推入地面和直根抵制连根拔起。迎风支撑,在地上细根,容易脱离。(1 c)作为测试所得背风支持最终失败,打破对支撑。旋转的中心(C.O.R.)变化,关于这个背风树旋转铰链和直根杠杆的地面或断裂。 (2a) Buttressed tree with sinker roots. The tree is anchored into the ground by the thick buttressed laterals by their sinker roots and by the tap-root. As the tree is pulled over material in the base of the trunk. In some species, the trunk has a narrower diameter beneath the top of the buttresses than above them. Presumably the transmission of stresses through the buttresses allows the tree to remain mechanically viable with less material in the lower butt. This can be seen at its most extreme in species with highly developed stilt roots. These, we must presume, act like flying buttresses, transmitting stresses widely to the extensive lateral root system. In some stilt-rooted species the primary stem is extremely thin beneath the stilt roots. It is much too narrow to support the weight of the tree above. The development of stilt roots allows palms to gain more height for a given stem diameter than those without stilt roots (Schatz et al. 1985). Fisher (1982) found reaction wood in the stilt roots of Cecropia species, possibly indicating the requirement for a greater tensile strength in buttresses of more limited cross-sectional area. Stilt roots are not a common feature of trees of well-drained lowland rain forest. For instance, only 5 out of 246 species surveyed in Gabon, West Africa, had stilt roots (Reitsma 1988). Species of swamp forest are much more frequently stilt-rooted.
使用一个方法基于异速生长,Ennos (1993 b)已经表明,root-plate系统的机械效率与工厂规模增加,因为产生的安克雷奇-土板(很大程度上由其重量)增加速度比干强度大小。是有效的在一个大的树,主根需要同样尺寸的箱子,甚至更大。在
(2 b),树旋转点只是背风的一面。最初的根牢牢锚树在地上,背风侧抵制被推入地面和直根抵制连根拔起。迎风支撑,在地上安全地举行的伸卡球根,还经受连根拔起,而是开始脱层。随着测试的进行(2 c)背风支持最终失败,打破最后的支撑。旋转中心的变化,关于这个背风树旋转铰链和迎风根继续脱层。(3)Non-buttressed树(Mallotus wrayi)。树是固定在地上的主根和一定程度上的外侧的根源。(3 b)树开到路边树绕点旋转背风一侧的主根的深度ca。0.5。背风侧根稍微推到土壤,然后扣而侧根迎风一侧抗拒被拉起来,表演张力。直根推到土壤背风一侧略微弯曲和旋转上面的中心旋转和下面这个弯曲,动作稍微迎风。 A crevice is formed on the windward side as the tap-root rotates. As the test proceeds (3c) these root movements continue, the leeward laterals buckling, the windward laterals uprooting and the tap-root pushing into the soil, increasing the size of the crevice. After Crook etal. (1997).
热带土壤,树可能难以产生大主根到足够的深度,需要足够的锚固,尤其是在物种大量木材。骗子& Ennos(1998)分析了机械故障的直根Mallotus wrayi树木在沙巴丹浓谷。在较小的树木树干断裂严重加载时通过绞车,但在更大的个人失败是通过tap-roots流离失所。这表明,大树不能产生足够大的tap-roots锚自己有效。
总之,现在有证据表明,拱结构,有效发挥重要作用在支持和锚定大树。然而,许多问题仍有待回答。在森林里做许多unbuttressed大树都有大tap-roots ?拱和root-plate系统具有成本效益的材料需要构建他们吗?做拱执行其他角色,如在森林的地面上占用空间减少的可能性建立竞争树?
靠树
大多数树木直立,甚至倾斜的地面上。这是因为它是机械更有效的支持下直接冠并保持主应力线内的树干,树干基地。此外,倚树获得高度在一个更大的施工单位成本高,这将导致一个高度增长的速度较慢。Loehle(1986)指出,大多数的树木做精益很小,与早期快速增长的物种繁殖或瘦成永久的高亮度条件如河流。倚在小树的成本低于大公司,因为他们可以利用木材的弹性性质与一些额外的支持精益投资支持。大树需要支撑较粗的树树干和更大规模的树基地精益。这些代表了重大投资,才会有价值倾向的回报很高。快速增长的物种可能通过阴影死亡风险如果不瘦到附近的缺口。也有可能,他们的低密度木材使倾斜更少的问题。Ishii &东(1997)获得了类似的结论比较小和大树生长在山坡上的梯度相对光照垂直于斜坡。 The quantitative model they derived has been criticised as unrealistic (Loehle 1997), even though its general predictions were found to be correct for a comparison of two species growing on slopes in warm temperate rain forest in Japan. None of these hypotheses have been explicitly tested for tropical雨林的树木。倾斜的河岸树木已经指出。例如,Dipterocar-pus oblongifolius特点是银行的一些河流在马来半岛和婆罗洲,习惯性地倾斜,经常在河里(Whit-more 1975)。
树下降
尽管树木生长的能力,减少局部高应力强度的可能性,机械故障是常见的热带雨林。在巴拿马Barro科罗拉多岛,310年一项调查倒下的树木(磨蹭et al . 1983年)发现,70%的树木折断的树干,25%已经被连根拔起,5%已经破掉在地面上。树木连根拔起往往比了,树干的直径更大的对于一个给定的高度,和密度,硬和更强的木头。支撑的程度似乎并未与倾向于吸附或拔出。易感性飓风灾害被发现与木材密度是负相关的树种在波多黎各(J.K.齐默尔曼et al . 1994年)。然而,对于5种从考艾岛,夏威夷,阀杆弯曲弹性模量是一个更好的关联飓风期间拍摄的可能性比木材密度(Asner &戈德斯坦1997)。僵硬的茎较低的弹性模量更有可能生存飓风不会断裂。木材的力学性能的影响一个物种的能力成功地抵抗风荷载和其他外部压力,但还需要更多的研究来确定哪些属性是最有影响力的。
它不能假定倒下的树木总是死。10厘米的165棵树胸径及以上,在1976 - 1980年期间Barro科罗拉多岛,88年(53%)re-sprouted从破碎的基地,其中26例(16%)在1987年仍然活着(磨蹭&布罗考1989)。82年巴西大西洋森林,100年树木连根拔起了新的芽,主要来自树干的底部(Negrelle 1995)。在这两项研究指出,在re-sprouting先锋物种很穷。笨蛋&布罗考(1989)发现,较小的树更有可能re-sprout长期成功。Negrelle(1995)指出,Tapirira guianensis尤其擅长连根拔后再生,并能从整个re-sprout树干的长度。
Sap上升
大自然的奇迹之一是大树的明显缓解大量的水提升到高处时发生。需要大量的水树顶,不仅取代水期间不可避免地失去了在光合作用吸收大气中的二氧化碳,还因为蒸发损失的水是一个非常有效的方式冷却叶片暴露在强烈的热带阳光。通过tiny-diameter树木质部汁液提升管道由列相似或容器元素。对重力提升的驱动力是自由能之间的差异在大气中气态水的叶子和他们的世界讲述表面和根的液态水。木质部汁液上升的普遍为人们所接受的内聚压力的理论提出的水的蒸发从叶子的细胞壁产生微观半月板产生很强的紧张局势在连续列的水从这些网站的根木质部导管。水的粘性特性允许列下张力拉而不是休息,因此提供蒸腾流。cohesion-tension理论预言存在的紧张局势应该在木质部中找到。至少1条(0.1 MPa)需要克服重力对于每个增量高10米,和等量可能是要求计数器的内部管道的流动阻力。
测量由Scholander压力炸弹被视为cohesion-tension的辩护理论。气体压力在炸弹来表达的sap测试拍摄假定等于木质部紧张,和测量值通常下跌cohesion-tension理论预测的范围之内。然而,炸弹不直接测量木质部紧张的压力。可以构建一个探针与一分钟压力传感器,可以正确插入血管测量内部压力。这些设备一直未能记录大植物芽的紧张局势(齐默尔曼et al . 1994年),包括在热带雨林的树高。其他反对cohesion-tension理论包括理论和实验研究水的列,似乎不太可能,预测的紧张局势可能存在于木质部(史密斯1994)。cohesion-tension理论的支持者发起了一系列的反驳和实验演示支持该理论的继续支持。压力传感器方法容易受到技术持怀疑态度。探针尖比孔小的管道被测量,和技术要求上的洞血管壁由探针海豹的条目完全没有管道中的压力改变,也没有空气进入,引起栓塞。液体探测器容易空化,因为气体种子内部表面(魏et al . 1999年)。 Milburn (1996) has also pointed out that the probe only accesses the surface vessels. These are liable to have lower tensions than average, either because they are still-living proto-xylem or because water recycling from phloem transport, so-called Munch water, rehydrates the outermost vessels. Others have used centrifugation techniques to show that significantly negative water pressures exist and can be maintained in xylem and that they correlate well with pressure-bomb measurements (Holbrook et al. 1995; Pockman et al. 1995). They also argue that glass capillaries used in experimental studies of water columns and their cohesiveness may not be realistic analogues of xylem conduits.
最近,精明的(1997 a, b)使用快速冷冻技术来想象向日葵叶柄木质部导管的活动。本研究的令人惊讶的结果是频繁的空化的证据以相对较低的紧张和快速re-filling船舶运动的水从管道周围的薄壁组织细胞。血管形成空洞的比例在一天不配合木质部张力的变化以一个压力炸弹:血管较少形成空洞当炸弹平衡压力是最大的。这是与经典cohesion-tension方差理论。精明的支持compensating-pressure木质部函数理论(精明的1995)。这个建议木质部运作比cohesion-tension理论预测以低得多的紧张局势,因为从周围的细胞,补充正压例如,石碑,射线薄壁组织或韧皮部,允许这是可能的。此外,标志着能力填写成泡渠道意味着水供应紧张时可以由木质部超过正常低操作值。compensating-pressure理论反过来已经会见了怀疑。其配方不符合公认的模型生物物理学(康斯托克1999)。死亡是没有显示的差异容易栓塞,预期的补偿压力从附近生活组织需要保持木质部内的流动(斯蒂勒和斯佩里1999)。
总之,最近怀疑促使研究人员生产强cohesion-tension理论验证。
树水力学
技术准确测量液流的速度在树木和植物的内部阻力流动现在可用。使用速度补偿热脉冲技术,贝克尔(1996)发现,个别树木强烈的白天的液流速率,皇冠区域线性相关,与不同的物种,和不同的网站的不同森林类型,安装相同的回归(图2.10)。之间的回归斜率改变湿和干燥的季节。同样,安德拉德et al。(1998)发现,树的大小是树形用水的主要决定因素在巴拿马,35米个人Anacardium excelsum估计每天使用379升。液流率与入射太阳辐射,这是蒸腾的驱动力(图2.11)。
是提供一个可以利用的水店当水蒸发蒸腾的需求很高。水店就会成指数级增长,树的大小尺寸和用水的体积从干细胞存储可以代表9 - 15%的每日总用水量(Goldstein et al . 1998年)。阀杆与水时期低transpirational充电的需求。
热带树种表现出广泛的不同措施的渗透系数的值可以估计(批&大口水壶1996)。一般来说,热带树木平均抑制电导相比,高值温带树木和裸子植物。快速增长,soft-wooded物种通常leaf-specific电导率值最高,即抑制电导单位叶面积测量段远端。因此他们应该能够快速提供大量的水每片叶子和维持高水平的蒸散在相对较低的水势梯度。这是实现不是由单位边材截面高导电性,但通过一个高比例的边材截面的分支。
branch-specific叶面积的边材横截面积比(LA / SA)似乎是分支蒸腾速率的一个重要的决定因素。四个不同物种的个体不同大小的增长在巴拿马之后基本相同的电导和蒸发的需求之间的关系当了由拉/ SA(·美茵茨合作撰写et al . 1997年)。换句话说,蒸腾速率单位叶面积时相同的LA / SA被允许,因此洛杉矶/ SA更具影响力蒸腾比气孔导度在许多情况下,因为通过低边界的解耦
-

- 0 100 200 300

图2.10关系意味着每天白天的液流和预计皇冠区域dipterocarp-forest树(三角形)和石南林树(圆圈)在湿(固体)和干燥(打开)时期在文莱。减少主轴回归斜坡湿和干燥的时间明显不同。后,贝克(1996)。
0 600 1200 1800 2400
当地时间
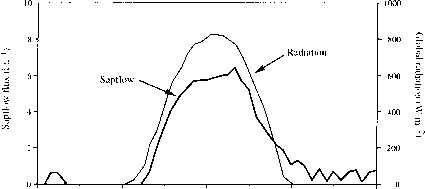
图2.11的日常课程短波辐射和液流Neonauclea树一个阳光明媚的天至少在旱季(喀拉喀托火山),印度尼西亚。后Bruijnzeel &学监(1995)。
0 600 1200 1800 2400
当地时间
图2.11每日的短波辐射和液流Neonauclea树在至少一个阳光明媚的天在旱季(喀拉喀托火山),印度尼西亚。后Bruijnzeel &学监(1995)。
层导,特别是在静止空气条件。
热带雨林物种似乎极易木质部栓塞,尤其是在比较与季节性干旱森林物种(批et al . 1998年)。比较物种混合龙脑香科树森林和森林健康从文莱没有显示任何显著差异在栓塞脆弱性之间的网站(批et al . 1998年)。自由排水的土壤健康的森林可能会选择与脆弱的木质部低于龙脑香科树森林物种。
树叶的液压电导的热带树种被发现跨越温带物种类似的范围所示(批et al . 1999年)。测量值可能足够低,表明,在一些物种中,叶提供了最大的耐水性植物连续的运动。比叶面积与水力导度显示没有明显的关系。
使用估计的木质部液流率和营养浓度,巴克和贝克(1995)能够计算营养的交付率的冠Dryobalanops aromatica树木在文莱。木质部汁液浓度的重要元素被发现,氮>钾> Ca > p养分浓度没有关系树的大小,并且经常显示与液流率正相关。交付率/树达到约40更易与h“我为N, 30更易与h”我K, 5更易与h“我为Ca和3 mmolh_1 P。
树结构
的形式建设的地上部分茎和分支的树中的一般模式在空间由两个因素:遗传bauplan拍摄的生长和分支和芽的命运与环境的交互。哈莉·& Oldeman(1970, 1975)提出了一系列23建筑模型树的形式,每一位植物学家命名的植物形态学的研究作出了贡献。这个系统是空中的类型学基因控制组件的轴组织基于角色状态的主要轴,分支,休息的时间在开发中,相对的花序顶端优势和立场。最近,罗宾逊(1996)利用符号逻辑来表示和分析哈雷& Oldeman系统。他推断出可能存在的几个,到目前为止,未报告的模型。
许多树在森林里似乎不符合一个23基本模型,即使考虑分支由于损伤或疾病的损失。这未能符合通常是由于这一过程哈莉·& Oldeman称为重复(哈莉·et al . 1978年)。一个或多个分生组织,而不是进行与原模型在适当的地方,重新开始,有时与一个完全不同的模型。重复似乎树形式发展的一个重要过程。它允许树大架构的灵活性。
一些树从来没有重申但符合单一模型在他们的生活。大树,哈雷(1986)把这称为巨人症,指出三组发生这种情况很普遍。这些都是松柏(例如南洋杉spp。),被子植物的家庭通常被认为是相对原始(例如肉豆蔻科)和快速增长,soft-wooded Cecropia物种等热带树木。所有的团体使用简单,稀疏支冠用树叶来填补空间。松柏,长寿树叶坚持远回树枝让他们高效聚光单元。在快速增长的物种,巨大的叶子在树枝的末端产生一个皇冠的外壳的叶子一层或两层支持相对较低的投资木材。这些热带遮阳伞树可以被认为是性早熟的:繁殖的“少年”建筑阶段。
建筑模型是一个有用的工具在热带树木和有一个清晰的描述系统组件(凯勒1994),但迄今为止进展相对较少涉及特定的模型与特定的生态角色(Bongers & Sterck 1998)。这可能是因为字符用于定义模型不够精确反映任何生态皇冠形式的有意义的人际关系。两种相同的架构模型在森林里可以看起来很不同,和另外两种不同的模型可以非常相似。
这并不意味着树形态不是与生态有关。分支模式影响叶的显示,机械效率支持和叶的水供应。所有这些因素可能作为选择压力皇冠的演变形式。比较两种treelet秘鲁低地雨林表示皇冠形式的重要性(Terborgh &马修斯1999)。考点chlorantha被发现在网站更直接的开销照明平均比Rinorea viridifolia,一般树冠的小缺口。教育部考试中心更高度支化的壳下垂树叶以外的皇冠。Rinorea有轮生的分支与平面层枝叶数组。后者设计更有效地拦截出现大量的横向光森林火情的网站。
异速生长
有机体的相对尺寸的变化,因为它的增长,或一系列大小不同的生物体,可以反映出一系列的约束与增长。这些可以物理尺度部队需要按比例少,甚至更多,投资在某些器官如果生物体维持一个统一的可能性有关损坏或破坏的力量。或者,可能反映了不同部位的大小相关的投资尺度在每个成长阶段进化压力优化性能。异速生长的研究也有用实证工具来洞察某些措施的信心可以用作其他代理。例如,许多个人和站特征往往估计从树树干直径测量。
最经常研究异速生长的关系树的茎直径(D)和高(H)。一个主要目标是评估异速生长的常数的大小,D oc哈,并确定其近似预测值基于工程或其他原则。之间的线性关系(= 1)两个异速生长的变量被称为几何相似。统一的锥度和材料组成的独立的列需要它的直径增加的速度比高为了维持一个恒定的机械稳定性。工程预计一个= 1.5弹性相似条件。研究高直径关系的热带树木的高度范围,对个别物种或样本大型化,表明弹性相似高直径关系可能是一个合理的近似(富et al . 1986;王1991;O ' brien et al . 1995),如图2.12所示。这意味着小树相对比大旧细长的鼻子。(这是一个事实,大多数人都有意识地不知道还用下意识地确定相对的树的高度当没有对象可用于视图。)
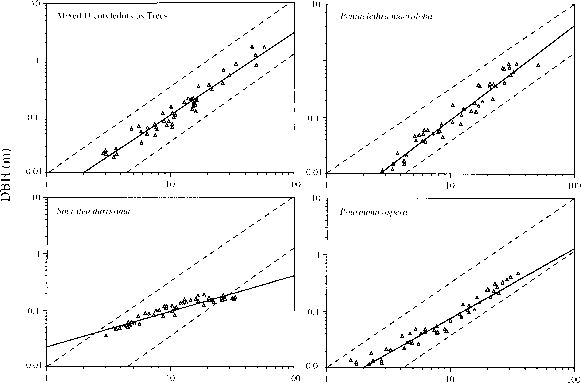
图2.12异速生长的茎直径(胸径)和高度混合双子叶的树,树冠主导树Pentaclethra macroloba,粗Pourouma gap-dependent树状棕榈Socratea durissima哥斯达黎加的热带雨林。p . macroloba包含20%的个人。实线是一个线性回归。上面的虚线是异速生长的曲线记录大小北美树木和较低的虚线理论屈曲极限的“平均”树,超出后树将扣在自己的weioht富π问H Qftfi ^
高度(米)
图2.12异速生长的茎直径(胸径)和高度混合双子叶的树,树冠主导树Pentaclethra macroloba
继续阅读:矿物营养
这篇文章有用吗?
读者的问题
-
法拉3个月前
- 回复
-
卢卡斯4个月前
- 回复
-
天使穆尔5个月前
- 回复