气孔导度
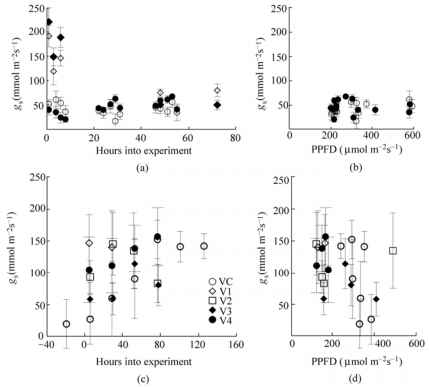
2004年对照植物V2期叶片的gs测量值显著高于2007年R2 - R4期(图16.2(a)),但与2008年对照植物V2期叶片测量值相当(图16.2(c))。埃塞克斯叶片VC期的gs值随着实验时间的推移不断增加,其他阶段的gs值在实验时间内没有变化趋势(图16.2(c))。2004年和2008年,Williams 82和Essex对照植物的gs测量对PPFD相对不敏感(图16.2(b))。2007年埃塞克斯对照植物的gs测量对PPFD同样相对不敏感,但在PPFD的相同范围内显示了更大的gs范围(图16.2(d))。即使2007年在每个工厂的白天进行了4到5个g的测量,在测量中没有明显的昼夜周期性。测量的可变性限制了确定处理效果的能力,部分原因是叶片气孔开口的可变性。
在2004年的研究中,暴露第一天V2叶片上的gs测量值仅与对照显著不同(学生t检验,以下称为P = 0.10的“t检验”)(图16.3(a))。注意,紫外线照射植物的PPFD仅比对照植物低10 |imol m s - 40 ^mol m s。从紫外线照射3小时开始,暴露在紫外线下的埃塞克斯的g值比对照植物大,但并不显著。同样的,
0 40 80 120 I lours进入实验(c)
图16.2品种和叶节点对叶片气孔导度的影响。图(a)中R2-R4阶段(圆形表示2007年测量值)和V2阶段(菱形表示2004年测量值,方形表示2008年测量值)的Essex和Williams气孔导度。2007年对照叶的PPFD与气孔导度的关系见图(b)。填充符号表示Williams 82,开放符号表示Essex cv。图(c)表示了VC ~ V4(2008)发育阶段Essex叶的气孔导度,图(d)表示了2007年对照叶的PPFD与气孔导度的对应关系。误差条为差值的标准误差。所有测量均在温室实验中进行,仅在暴露6小时(第一天结束)后,暴露在紫外线下的Williams 82的g值显著低于对照植物(t检验P = 0.10)。在第一天之后,随着紫外线照射量的增加,紫外线照射对gs的影响逐渐减小(图16.3(a))。第三天测量的太阳辐照度(120 ^mol m-2s-1 -140 ^mol m-2s-1)低于前几天(120 |imol m-2s-1 - 200 ^mol m-2s-1),但似乎没有影响植物的反应。
在2007年的研究中,从繁殖R2到R4阶段,紫外线照射的埃塞克斯叶的许多gs测量值与对照植物相比略有较大,但差异不显著(t检验P = 0.10),主要发生在实验进入31小时后(暴露第二天结束)。增加
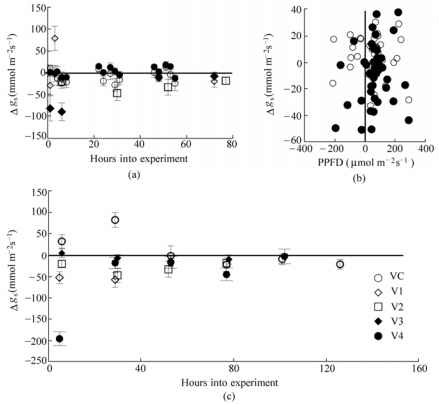
图16.3紫外线照射对叶片气孔导度的影响。R2-R4阶段(圆形表示2007年的测量值)和V2阶段(菱形表示2004年的测量值,方形表示2008年的测量值)的Essex和Williams气孔导度。2007年UV叶片与对照叶片的PPFD差异与气孔导度的关系见图(b)。填充符号表示Williams 82,开放符号表示Essex cv。图(c)显示了埃塞克斯叶在VC到V4发育阶段(2008)的气孔导度。差异用UV-Control表示。误差条是标准误差的差值。所有测量均在温室实验中进行
图16.3紫外线照射对叶片气孔导度的影响。R2-R4阶段(圆形表示2007年的测量值)和V2阶段(菱形表示2004年的测量值,方形表示2008年的测量值)的Essex和Williams气孔导度。2007年UV叶片与对照叶片的PPFD差异与气孔导度的关系见图(b)。填充符号表示Williams 82,开放符号表示Essex cv。图(c)显示了埃塞克斯叶在VC到V4发育阶段(2008)的气孔导度。差异用UV-Control表示。误差条是标准误差的差值。所有测量均在温室实验中进行,在R2至R4期间,与Williams 82叶片紫外线暴露相关的gs在P = 0.10时不显著。作为所有测量小时的汇总样本,紫外线暴露导致Williams 82在R2至R4范围内的gs有统计学意义上的下降(t检验P = 0.05),但在Essex没有统计学意义上的增加。在2008年的研究中,紫外线照射的埃塞克斯植物在VC期的前2天比对照有较大的gs (t检验P = 0.10)趋势(对应于对照叶片的低gs值);然而,到第3天,暴露叶片与未暴露叶片之间的差异消失(图16.3(c))。在随后的所有营养阶段(从V1到V4),紫外线照射下的埃塞克斯叶片的g值与对照相同或增加(图16.3(c))。 As a pooled sample of all hours of measurement, the UV exposure resulted in a statistically significant increase in gs at VC (t-test P = 0.01) and statistically significant decreases at V1, V2, and V4 (t-test P = 0.01), but not at V3.
在2004年的两项田间研究中,V5的uv暴露植株和对照植株之间的gs无显著差异(表16.2)。尽管这样的结果可以预期,因为温室实验表明,随着植物增加紫外线照射,紫外线照射对gs的影响呈下降趋势(图16.3(a)),但重复中测量的gs的广泛变异性导致无法得出明确的结论。
表16.2 2004年实地研究结果
UV-Control
V5阶段
R3阶段
表16.2 2004年实地研究结果
UV-Control
V5阶段
R3阶段
品种 |
测量 |
的意思是 |
SE |
的意思是 |
SE |
埃塞克斯 |
Gs (mmol m2’) |
-140年 |
158.1 |
52.9 |
187.5 |
E (mmol m2’) |
-1.35 |
0.558 |
0.859 |
0.830 |
|
A (pmol m2 s’) |
- 2.33 * |
1.28 |
-1.15 |
2.25 |
|
WUE (x 10 4) |
2.28 |
3.29 |
- 4.12 |
3.07 |
|
威廉姆斯82 |
Gs (mmol m2 s 1) |
- 23.6 |
142 |
322 |
367 |
E (mmol m2 s 1) |
-1.44 |
0.863 |
0.395 |
1.06 |
|
A (pmol m2 s 1) |
- 2.07 |
1.91 |
0.204 |
1.21 |
|
WUE(x 10 4) |
3.89 |
2.95 |
0.0268 |
2.39 |
* = P = 0.10,差异有统计学意义。
* = P = 0.10,差异有统计学意义。
总的来说,紫外线照射对埃塞克斯的影响对应于植物暴露开始时(V1到V4和R2到R4) gs的增加,随着暴露时间的增加在一定程度上得到补偿。这与V5和R3的gs无显著差异的现场研究结果一致。Williams 82在V2和R2-4的初始表现为gs下降,在整个温室暴露期间变为不显著(P = 0.1时的t检验)增加。现场研究表明,V5和R3的gs变化可以忽略不计。一些研究表明,紫外线- b辐射会降低gs (Middleton and Teramura, 1993;Dai et al., 1995),而其他人则揭示了gs的增加(Musil和Wand, 1993)。Eisinger et al.(2000)研究表明,紫外线照射导致蚕豆气孔开放的两个主要峰值分别为280 (UV- b) nm和360 (UV- a) nm。Negash(1987)的研究表明,255 - 295 nm的紫外线辐射导致Eragrostis tef在紫外线照射15和45分钟后气孔迅速关闭。Battaglia和Brennan(2000)在黄瓜子叶(Cucumis sativus L. cv。而在向日葵(Helianthus annuus L cv。 Gray Stripe). Some studies suggest that the gs in plants vary greatly because of the difference in their light requirement
(Kozlowski and Pallardy, 1997)。2004年、2007年和2008年的温室研究表明,没有证据表明PPFD对gs的影响(图16.2(b), (d))。不同品种间接受UV- b量的差异,Essex接受的UV- b暴露量比Williams 82多7%,可能部分导致了本研究中发现的植物反应差异,并且可能是长时间UV- b暴露的重要因素。
气孔导度通常表现出类似于保护细胞长度减少的响应模式(giz et al., 2005)。Nogues等人(1998)发现暴露在高水平UV-B下的豌豆(Pisum sativum)电导的降低主要是保护细胞改变的结果。我们已经证明紫外线导致Williams 82的守卫细胞长度增加,而在Essex观察到守卫细胞长度减少(未发表的数据)。观察到的守卫细胞长度的改变是否由紫外线的伤害气孔首字母的发育或发育过程的结果,如Essex和Williams 82表现出不同的形态响应。
继续阅读:蒸腾作用
这篇文章有用吗?